
Africká savana
Savana
Savany zabírají přibližně 40% afrického kontinentu. Tvoří přechod mezi pouští a tropickými lesy. Přesto toto vymezení není dokonalé a dokonce není do současnosti přesně vědecky ustálené, podobně jako pojem les. Savana by se dala popsat též jako bojová zóna mezi stromy a travami. Nezanedbatelným činitelem je oheň, který nastoluje určitou rovnováhu v tomto ekosystému. Na rostlinnou složku se váže i množství živočišných druhů.
Pokud se chcete dozvědět více, přečtěte si následující text – k navigaci slouží „Obsah“ …
(text a obrazový doprovod jsou výsledkem diplomové práce autora stránek, vypracované na Přírodovědecké fakultě UK v roce 1999, pod názvem „Africké savany“).
OBSAH
1. ÚVOD
1.1. Cíl práce,informační zdroje a metody zpracování
2. Definice savany
2.1. Přírodní nebo antropogenní původ savan
2.2. Strukturní a funkční charakteristiky savany
2.1.1. Fyziognomická charakteristika
2.1.2. Sezónní změny v savanách
2.1.3. Množství a změny v primární produkci
2.1.4. Kvalita primární produkce
2.1.5. Dynamika ekosystému savany
2.1.6. Souhrnná charakterizace savan
3. Geografické rozšíření savan ve světě
4. Africké savany
4.1. Fyzicko-geografické podmínky
4.1.1. Klimatické podmínky
4.1.1.1. Sluneční záření – solární radiace
4.1.1.2. Srážky
4.1.1.3. Teplota
4.1.1.4. Výpar
4.1.1.5. Klasifikace klimatu
4.1.2. Geologické a pedologické podmínky
4.2. Typologie savan
4.3. Vegetace africké savany
4.3.1. Stručná charakteristika
4.3.2. Flora africké savany
4.3.2.1. Floristická příbuznost a fytogeografie
4.3.2.2. Geografické rozšíření flóry Afriky
4.3.2.3. Životní adaptace
4.3.2.4. Morfologie savanových druhů rostlin
4.3.2.5. Životní formy
4.3.2.6. Kooperace mezi bylinnými a stromovými rostlinami
4.4. Hlavní vegetační kategorie savany
4.4.1. Uzavřené formace
4.4.2. Otevřené formace
4.5. Fauna
4.5.1. Zoogeografie
4.5.2. Bezobratlí
4.5.3. Obratlovci
4.5.4. Potravní vztahy
4.6. Oheň v savaně
4.7. Vliv člověka
4.8. Ochrana přírody a národní parky
1. Úvod
Africké savany se těší značné pozornosti mnoha přírodovědců a turistů z celého světa. Rozkládají se v širokém pásmu tropické oblasti, přerušeným klínem tropického deštného lesa ve středu a na západě kontinentu. Na jihu a severu jsou ohraničeny pouštními a polopouštními formacemi. Hlavním lákadlem tohoto biomu jsou velká savanová zvířata. Zájem veřejnosti je koncentrován převážně na ně. V důsledku dramatických změn s rozvojem lidské společnosti je potřeba porozumět vztahům a závislostem organismů v ekosystému savan a vyřešit tak ochranu přírody v afrických národních parcích.
Savany jsou pro většinu lidí jen holé pláně Serengeti, přestože většina typů savan má k této představě daleko. Jak se dnes předpokládá, biom savany je „válečná zóna“ mezi stromy na straně jedné a trávami na straně druhé. V určitých podmínkách mohou převládnout stromy a keře, potom savana vypadá jako „lesní“ ekosystém.
Smyslem práce je podat co nejúplnější charakteristiku biomu savan. Jsou zde zachyceny hlavní fyzicko-geografické podmínky ovlivňující rozšíření savan v Africe. Dále je zde pozornost věnována savanové vegetaci, na které jsou přímo či nepřímo závislí všichni živočichové. Činnost člověka je zde zachycena v roli ochránce i ničitele tohoto pro nás „exotického“ prostředí.
Důvodem proč jsem si vybral toto téma byl můj čtyř měsíční pobyt v oblasti savan s expedicí AFRICKÁ MOZAIKA 1997. Pestrost fauny a flóry a její sezónní proměny učarují každému, kdo tuto oblast navštíví.
V práci není zmiňován ostrov Madagaskar, ačkoli k Africe geograficky patří. Jiné fyzicko-geografické podmínky a značná časová izolovanost zde vytvořily zcela odlišné životní prostředí . Madagaskar vytváří přechodnou zónu mezi oblastí Afriky a Asie a zasloužil by si vlastní zpracování.
Celou magisterskou práci jsem napsal pomocí textového editoru MS WORD. Tabulky jsou zpracovány v programu EXCEL. Při úpravě map a jejich skenování mi pomáhali M. Krčma a V. Treml. Fotografie v textu zhotovil autor magisterské práce.
1.1. Cíl práce, informační zdroje a metody zpracování
Cíl práce
Cílem práce bylo popsat celkovou charakteristiku savan, zvláště Afriky. Zhodnotit fyzicko-geografické podmínky, nastínit základní ekologické vazby mezi organismy v tomto biomu, ale též zachytit činnost lidské společnosti v savanách. Role člověka v savaně je dnes natolik významná, že v některých místech nelze rozlišit jestli je savana přírodního nebo druhotného původu. Vliv člověka na savanu by si však zasloužil vlastní zpracování. Cílem práce je tedy podat celkový obraz o afrických savanách, jejich fyziognomii, proměně a též značných ekologických problémech. Celkovému obrazu by mělo napomoci i množství fotografií, map a tabulek.
Z uvedených cílů vyplývá struktura práce, která je rozdělena do čtyř tématických částí. V první části je definováno, co jsou savany a jaké jsou její základní charakteristiky. Pozornost je věnována i jejich rozšíření ve světě. Ve druhé části je kladen velký důraz na fyzicko-geografickou charakteristiku, vzhledem k tomu, že právě fyzicko-geografické podmínky vytváří pro výskyt savan rozhodující rámec. Třetí část popisuje vegetaci a faunu africké savany. Čtvrtá část je věnována člověku, který je hlavním modifikačním činitelem v rozšíření savan. Závěr pak shrnuje výsledky práce.
Prameny a literatura
Prameny a literaturu, ze kterých bylo čerpáno, můžeme rozdělit podle různých kritérií a přístupů k problematice. Při charakterizaci afrických savan bylo čerpáno z mnoha vědních disciplín. Daly by se proto rozdělit na díla biologická, geografická, sociologická, ekologická atd. Jiným kritériem může být povaha díla: atlasy, učebnice, časopisy, novinové články a jiné odborné a vědecké práce.
Tato práce si klade za cíl být syntetickou prací oborů geografie a biologie, proto bylo využito následujícího dělení:
a) Díla odborná zabývající se biologickou a ekologickou problematikou
Do této skupiny patří díla, zabývající se nejen problematikou ekologických vazeb, dále různé atlasy živých organismů, systematické učebnice a určovací klíče. K stěžejním dílům zde použitým patří ekologická publikace Ekosystems of the world – Tropical savanna, vydaná v zahraničí. Dalším cenným zdrojem byl časopis pro biologickou práci Živa. V ročnících 23. a 24. zde Prof. Ing. J. Jeník DrSc. popsal základní ekologické vazby v afrických savanách. Díla ze kterých byla čerpána a přebírána data, jsou průběžně uváděna v textu. V seznamu literatury jsou uvedena díla, která slouží k získání znalostí, seznámení s terminologií a pro základní orientaci v biologických oborech.
b) Díla odborná zabývající se fyzickou geografií
Rozhodující část literatury, která byla použita, vznikla na pracovištích Přírodovědecké fakulty Univerzity Karlovy v Praze. Přestože se žádné dílo nezabývá africkou savanou, patří Fyzická geografie I. a Fyzická geografie II. k nejdůležitějším zde použitým dílům. V časopise Geografické rozhledy lze v některých číslech najít články, týkající se Afriky a přeneseně i africké savany. Některé články i zde citované jsou od vedoucího této magisterské práce RNDr. Zdeňka Klimenta CSc.. Jednotlivá čísla Geografických rozhledů jsou jmenována v seznamu literatury.
c) Díla populárně vědecká
V současné době jsou velmi dostupná díla populárně vědecká, která přináší základní zdroje informací i o afrických savanách. Některá díla též poskytují i značně obsáhlé a podrobné informace týkající se ekologie, biologie a geografie afrických savan. Mezi hlavní populárně vědecké knižní práce patří díla Josefa Vágnera, bývalého ředitele zoologické zahrady ve Dvoře Královém. Velmi dobrou populárně vědeckou knihou je biologicko systematické dílo Obratlovci. Dále ještě jmenujme z edice Larousse dílo Savany a z Knižního klubu Atlas panenských míst. Naopak cenným zdrojem geografických informací je časopis National Geographic vydávaný National Geographic Society ve Washingtonu. Tyto a ostatní populárně vědecká díla jsou uvedena v seznamu literatury.
d) Mapová díla
Důležitým zdrojem informací jsou též mapová díla. Základní údaje zde použitých byly čerpány z map: Michelin, 1995, Afrique central et Sud, Madagaskar, Pneu Michelin a Půdní mapy světa FAO/UNESCO, 1974
Zvláštním zdrojem informací, které poskytuje základní pohled na ekologickou a biologickou problematiku ekosystémů, respektive afrických savan, je CD-Rom Encyklopedie přírody 2.0; Vlastníma očima. Na přípravě tohoto CD-Romu se autor magisterské práce aktivně podílel.
Metodika práce
Základní informace byly čerpány z výše uvedené literatury a pramenů. Všeobecně dostupná jsou populárně vědecká díla zabývající se jen některými aspekty biologie, geografie a ekologie savan. Je to pochopitelné, protože africké savany se těší velké oblibě mezi veřejností, díky populacím velkých savců. Vzhledem k tomu, že popisujeme celkovou charakterizaci savan, nejsou tyto informace dostačující. Výjimku představuje již zmíněná ekologická publikace Ekosystems of the world – Tropical savanna, která byla často jediným zdrojem informací a proto byly převzaty i citace z tohoto obsáhlého díla. I tato publikace v důsledku značné monotematičnosti, neposkytuje jiné než ekologické informace.
Důležité informace byly čerpány i z dobového tisku a periodik. Některé informace a údaje byly porovnány se samostatným šetřením a informacemi získanými při mé studijní cestě s expedicí AFRICKÁ MOZAIKA 1997.
Tato práce je prací kompilační, bylo proto pracováno s různými zdroji informací. Nemožnost provádět vlastní dlouhodobá odborná pozorování a měření je dána vzdáleností africké savany.
2. Definice savany
Definice slova „savana“ není běžně v přírodních vědách ustálena. To samé platí pro její francouzský (savane), anglický (savanna) a španělský (sabana) ekvivalent. Předpokládá se, že pochází ze starokaribského slova, ale jeho etymologie obsahuje nejasnosti. G.F. de Oviedo y Valdes ( 1535) určil, že slovo „sabana“ je možno vztáhnout na krajinu bez lesů s množstvím krátkých a vysokých trav. Ve třetím vydání Shorter Oxford Dictionary z roku 1944 je zaznamenáno, že slovo „savanna(h)“ je v Anglii známo od roku 1555 ze španělského slova „zavana“ , které označuje „pláně bez lesů“. Francouzský slovník slov z roku 1863-72 a Robert 1964-65 zmiňují používaní slova „savane“ od roku 1529 a jeho původ z karibského jazyka, ačkoli bylo interpretováno prostřednictvím španělštiny. Slovo savana je tedy kreolského původu. Nejvíce bylo tohoto termínu používáno kolonisty amerického kontinentu, od Kanady po Argentinu. Označovali jím krajinu, které se dnes říká prérie. Po dlouhou dobu se cestovatelé, botanici a přírodovědci drželi tohoto běžně rozšířeného slova.
První botanik, který užil v prezentaci slova savana v současném významu byl Grisebach (1872). Ve druhém vydání jeho Die Vegetation der Erde, se zmiňuje, že „savana se liší od mírných stepí zápojem stromovité vegetace“. Podobný pohled měl i Drude (1890), který se domníval, že “ savany nejsou tvořeny jen vysokou travou, ale též představují tropické stromovité rostliny, které opadávají během období dešťů“. Shimper (1898) svou definicí též míní, že „termínem louky jsou označovány travnaté oblasti s hydrofilními a tropofilními rostlinami. Když zde rostou xerofilní trávy, jsou to stepi a když zde roste xerofilní vegetace s izolovanými stromy, je to savana“.
Od této doby mnoho autorů používá termín savana označující společenstva mající plynulou travnatou vrstvu, obvykle s osamělými stromy. Beard (1953) definoval savanu jako – „začleněnou prakticky plynulou ekologicky dominantní vrstvu více či méně xeromorfních rostlin, ve kterých jsou trávy a ostřice hlavní složkou, s roztroušenými křovinami, stromy a palmami“.
Mnoho botaniků pracujících v Africe přijalo podobnou definici. Vědecký koncil pro Afriku na mítinku fytogeografie v Yangambii odsouhlasil formulaci savany jako: „Útvaru trav alespoň 80 cm vysokých, které tvoří dominantní vrstvu nižších trav, obvykle každoročně vypalovanou požáry. Trávy mají ploché listí, basální metabolismus a stonky. Obvykle jsou zde zastoupeny stromovité rostliny.“ ( Conseil Scientifique pour l´Afrique, 1956).
Společně s touto fyziognomickou kategorií, se na mítinku v Yangambii ustanovily ještě čtyři podkategorie savany. První jsou savanové lesy, tvořené stromy a keři se zápojem trav, které jsou obvykle světlé. Druhou je stromová savana s osamělými stromy a keři. Třetí je křovinatá savana s osamělými keři a poslední čtvrtou je travnatá savana, kde stromy nebo křoviny obvykle chybějí. Horské travnaté oblasti, zamokřené louky nebo pastviště byly zařazeny do samostatných skupin.
Z těchto definic vyplývá, že slovo „savana“ se používá pro bezlesou krajinu. Měla by však platit určitá kritéria pro tropické a subtropické savany: 1) trávy a ostřice tvoří souvislou a dominantní vrstvu, občas přerušeny zápojem stromů a keřů, 2) vyskytují se zde občasné požáry, 3) růst souvisí se střídáním období dešťů a sucha.
Většina současných fytogeografických a ekologických knih souhlasí s touto nebo podobnou definicí. Mnoho z nich též klade důraz na fakt, že z hlediska zonálního zeměpisu je savana ohraničena od „uzavřených“ tropických lesů po „otevřené“ suché stepi, s širokou přechodnou zónou od savanového lesa po travnatou step.
Přes všechna tato vymezení musíme být opatrní. Například východoafričtí autoři odmítají používat slovo savana mimo africký kontinent. Termín „savana“ se jeví jako nespolehlivý, protože „se vztahuje k celé stupnici různých biomů a má hodně synonym, používaných v různých částech intertropické zóny“ (Jeník, 1975). Ale z obecného pohledu, oba populární a vědecké protiklady – savana i les jsou běžné a budou nepochybně nadále využívány. V přehledu těchto rozdílných názorů na termín „savana“ není udivující, že existuje několik odlišných systémů klasifikace a nomenklatury pro tropickou vegetaci. Některé navrhované systémy jsou regionálního rámce. Jiné jsou univerzální. Současné představy různých základních systémů a názvosloví přineslo UNESCO, 1979.
2.1. Přírodní nebo antropogenní původ savan
Pro obyvatele tropických oblastí byla savanová krajina místem, které poskytovalo dobrý výhled do krajiny, nehrozilo zde přepadení dravci a byla možná spolupráce loveckých tlup. První obyvatelé zde snadno zakládaly pole a pastviny. Proto je obtížné přesně definovat vzniklou tropickou savanu za skutečnosti , že fyziognomické kategorie vegetace mohou být konečným výsledkem buď přírodních procesů nebo lidské činnosti, nebo kombinací obou dvou dohromady. Často je velmi obtížné zařadit dnešní savany do kategorie přírodní či antropogenní savany.
Vše začalo být velmi obtížně rozlišitelné s dnešním rozvojem lidské činnosti přispívající k značným změnám na plochách již existující savany. Situace je jednodušší v místech, kde ekosystém savany v minulosti neexistoval.
Přírodní „klimax“ savan existuje již po velmi dlouhou dobu. Rostliny (např. stín netolerující C4 rostliny ¤) a živočichové, kteří dnes žijí v savanách museli odněkud přijít a to buď z tropického lesa nebo suchých stepí a je velmi nepravděpodobné, že by se podmínkám savany přizpůsobili za dobu velmi krátké existence dnešního člověka. Příkladem jsou savany Jižní Ameriky, kde vliv člověka do současnosti je velmi malý nebo vůbec žádný.
Hlavní roli v ekosystému savan má množství fyzických faktorů – klimatických, pedologických i geologických. Mnoho autorů navíc podtrhuje význam deficitu vlhkosti během střídání krátkých vlhkých a dlouhých suchých období, malou úrodnost půd a půdně hydrologický režim. Dále pak sklon svahů, prohlubně, stupeň rozčlenění a přítomnost nebo nepřítomnost vrstvy laterických slepenců.
Člověkem vytvořené nebo podporované savany jsou zvláště dobře prezentovány v tropické Africe a Asii, kde se lidská aktivita projevovala a projevuje ve značném měřítku po celá tisíciletí. Požáry byly hlavním nástrojem pro odstraňování vegetace. Oheň je stále používán lovci v savaně k lovu – sběrači v Africe a v Austrálii. Též afričtí zemědělci a pastevci stále užívají ohně k získávání zemědělské půdy. (Klíma, 1970).
V mnoha částech západní Afriky byly savany vytvořeny člověkem a jsou stále udržovány záměrnými kontrolovanými požáry travin. To však neznamená, že požáry nemohou vzniknout přírodní cestou. Mohou být založeny blesky nebo samovznícením nakupeného materiálu. Tyto požáry jsou věrohodně doloženy z Afriky (Jeník, 1975). Dopad požárů na vegetaci je však málo intenzivní. V případě, že jsou některé plochy uměle udržované savany chráněny před požáry, rychle je pokryje během několika let listnatý les.
Převaha druhotných savan v mnoha částech Afriky přivádí mnoho ekologů k názoru, že na formování savan má vliv hlavně faktor antropologický, a to jak v původních savanách, tak v rozšiřování savan v jiných oblastech. Odlesňování, sběr palivového dříví a nadměrné spásání domestifikovaným skotem je důležitým faktorem a znamená větší destrukci než činnost ohně. Tyto vlivy byly pravděpodobně zdůrazněny hlavně klimatickými změnami v minulosti ( Aubréville, 1949, 1962).
Jinou teorií je teorie smíšená, která byla navozena některými ekology západní Afriky. Například, podle Adjanohoun (1963), současná savana v severním okraji guinejského lesního bloku a některé savany v tomto regionu jsou stále obklopeny deštným pralesem. Nemohou být tedy výsledkem dřívější činnosti člověka. Podle této teorie jsou pozůstatkem poslední suché fáze pleistocénu. Severní výběžky deštného lesa ustoupily jižně a savany tak dosáhly mnoha míst Guinejského zálivu. Člověk vstoupil do těchto míst mnohem později a svým působením udržoval otevřenou savanu při pozdějším návratu deštného lesa. Tato teorie působí přijatelně, ale je potřeba ji doložit lepší palynologickou, pedologickou a archeologickou dokumentací.
2.2. Strukturní a funkční charakteristiky savany
Savany sdílí ve všech částech světa množství strukturních a funkčních charakteristik, které je odlišují od ostatních travnatých terestrických ekosystémů tropických oblastí. Savany se formují v přechodné zóně mezi uzavřenými deštnými lesy po otevřené suché stepi. Jejich fyziognomická struktura se mění s rostoucí ariditou. Primární produkce má zřetelně sezónní charakter s kvalitativními a kvantitativními charakteristikami. Mají relativně jednoduchou strukturu a odchylky populace jednoho dominantního druhu mohou mít dalekosáhlý vliv na jednotlivé složky ekosystému.
2.2.1. Fyziognomická charakteristika
Fyziognomický gradient se postupně mění s gradientem klimatickým, s růstem ročních srážek roste i hustota stromů. Tuto závislost však narušují topografické rysy krajiny, působení ohně, býložravci a člověk. Fyziognomický gradient nemá z těchto důvodů lineární průběh.
V ekosystému savan spolu koexistují stromy a trávy a na mnoha plochách je mezi nimi stálá rovnováha. Tyto dvě životní formy, které jsou v jiných biomech antagonisté, mají jeden z druhého prospěch (Jung, 1969; Gerakis a Tsangarakis, 1970). Například v Fété Olé v Senegalu jsou pro fixaci dusíku mnohem důležitější traviny než stromy Acacia senegal a dokonce Balanites aegytiaca (Bernhard-Reversat, 1977). Naproti tomu, po suchu v letech 1972 až 1973 v Sahelu, byl návrat bylinné vegetace mnohem rychlejší v místech, kde pastevci nekáceli stromy a křoviny, nebo je neničili jejich stáda skotu a koz.
Neustalé hledání rovnováhy mezi dřevinami a travinami je jedním z nejzajímavějších hledisek ve vývoji ekosystému savan. Otázka, proč nepřevládne jeden z partnerů tohoto soužití není ještě plně vědecky objasněna. V neotropické oblasti jsou zřetelně patrné oblasti, které oscilovaly mezi deštnými lesy a savanami po celé období kvartéru. Savany se periodicky rozšiřovaly do deštného lesa, který zažíval epochy expanze a úbytku. Tyto opakované změny mohly být důvodem schopnosti stromů vytvořit vhodné podmínky pro šíření rostlin, vzniku humusu a půdy v látkovém cyklu savany.
Analýzou kompetice mezi dřevinami a trávami by se mohlo zdát, že stromová složka má hlavní vliv v ekosystému savan (Noy – Meir, 1979). Podle Noy a Meira způsobuje silný tlak býložravců vyhynutí vytrvalých trav a mění savanu v les. Dále pak stromy soutěží s travami o povrchovou půdní vrstvu a navíc využívají vodu uloženou hluboko pod povrchem. A tak trávy usychají v dostatečně husté stromové vegetaci. Stromy, na druhé straně, mohou přežít i v nejhustších porostech trav, jestliže srážky dostatečně nasytily i hlubší podpovrchovou půdní vodu a není pro ně problém rozšířit semenáčky.
Naopak profesor Jeník (Jeník, 1975) spatřuje jako dominantní životní formu početné druhy trav a šáchorovitých. Podle něj mají obě složky odlišné schopnosti, jak využívat potravní zdroje v prostředí a jakým způsobem přežívají nepříznivá období. Dřeviny potřebují trvalejší zásobování vodou, hůře snášejí větší výkyvy teplot a potřebují větší objemy půdy, protože jejich kořeny pracují extenzívně. Naopak traviny našly v savanách nejvyšší stupeň rozvoje a přizpůsobení. Vývojově řečeno, traviny jsou životní forma mnohem mladší a není vyloučeno, že dnešní přirozené savany jsou vlastně bojovou zónou mezi suchomilnými lesními stromy a expandujícími se a neustále zdokonalujícími travami. Tato nepříliš stabilizovaná rovnováha je někdy zvrácena ve prospěch otužilejších a lépe přizpůsobených trav. Všechny méně příznivé faktory, jako jsou nenadálé výkyvy teplot a srážek, mělčí půda na lateritických slepencích, okus velkých savců, podporují šíření trav. V přírodním systému, kde v blízkosti sebe žijí tak úspěšné životní formy, vznikají mnohé mezery a vakua v konkurenčním tlaku, které naopak pohotově zaujímají dřeviny, například trhliny v lateritických slepencích, místa s prosakující vodou, místa lokálně zúrodněná vlivem termitů nebo hlodavců atd. V celkovém součtu těchto mezer může pak vzniknout zdánlivě homogenní útvar stromovité nebo keřovité savany.
2.2.2. Sezónní změny v savanách
Sezónní variabilita klimatu a životních cyklů se odehrává ve všech ekosystémech, dokonce i v tak stabilním ekosystému jako je tropický deštný les. Sezónní změny v tropických savanách jsou zřetelné a podobné střídání ročních období v mírných šířkách. Savany nejsou pouze přizpůsobeny přežít dočasnou změnu, ale pravděpodobně požadují takovéto změny k udržení zdárného rozvoje, například semena hřebenatky trojmužné Dactyloctenium aegyptium (Jeník, 1975).
Sezonalita v cyklech savanových rostlin a živočichů se stává více zřetelnou, jak postupujeme od hranice tropického deštného lesa směrem k pouštím a polopouštím. Podle stejného gradientu se klimatické podmínky stávají více a více nepředvídatelné. Jsou zde velké každoroční výkyvy v množstvích, v časových změnách srážek a výskytu dlouhých období sucha. Pro přežití v takovýchto podmínkách mají organismy savan řadu morfologických, fyziologických a životních přizpůsobení.
Sezónnost organické produkce vede jen k dočasným potravním zásobám, které jsou využívány nomádickými sekundárními konzumenty a migrujícími živočichy. Dalším rysem je, že velká část živé rostlinné biomasy se nalézá pod zemí. V savanách Sahelu v Fété Olé na severu Senegalu je maximální podíl sušiny biomasy trav okolo 1.2t ha-1 nad zemí s porovnáním 1.48t ha-1 pod zemí. Mnohem dále na jih, v Lamto v parkové lesnaté savaně v Pobřeží Slonoviny je průměrný úhrn nadzemní rostlinné biomasy v rozmezí 3.2 až 4.36t ha-1 , kdežto podzemní biomasa dosahuje 10.1 až 19 t ha-1. Zatímco v Lamto činí podíl kořenové biomasy 28.9 až 31.8% z podílu sušiny biomasy, ve Fété Olé může dosáhnout až 43.7% (Lamotte,1978; Pandeya a Jain, 1979).
2.2.3. Množství a změny v primární produkci
Tropické savany jsou charakterizovány relativně vysokým množstvím čisté primární produkce spojené s tvorbou biomasy. Množství primární produkce by mohlo být větší, kdyby voda a úrodnost půdy nebyla limitována přírodními podmínkami. Primární produkce v savanách může občas překonat lesy rostoucí v podobných podmínkách. Například: čistá primární produkce, nad i pod zemí, bylinných a stromových druhů v Lamto v Pobřeží Slonoviny dosahuje 21,5 až 35,8 t ha-1 za rok (César a Menaut, 1974).
Většina savanové vegetace se dožívá velmi krátkého věku. Netýká se to pouze bylinných druhů, které vytvářejí velké procento nadzemní části biomasy (až okolo 85%) , ale též stromů. Mimo baobabů (několik stovek let) se velmi málo stromů a keřů v savaně dožije několika desetiletí. Například akácie se nedožívají více než 30 – 35 let, a pouze Balanites aegyptiaca a Grewia bicolor se občas dožijí okolo sta let (Poupon,1976).
Rychlá obměna populací živočichů malých velikostí je další charakteristikou savany. Mnoho ještěrů, hlodavců a snovačů se nedožívá průměrně ani jednoho roku. Jedna z největších mortalit mezi dospělci je pozorována u ptáků, například u Lagonostika senegala je až 70 % za rok (Morel,1969). Proto je potřebná vysoká fekundita k udržení rovnováhy populace, při tak velké mortalitě. Většina rostlin a živočichů savany tedy patří mezi čisté r-stratégy ¤¤.
2.2.4. Kvalita primární produkce
Z kvalitativního pohledu je většina primární produkce v savanách využita jako potrava sekundárních konzumentů. Jeden z hlavních rozdílů mezi ekosystémem tropického lesa a ekosystémem savan leží v rozdílných proporcích jedlé a nejedlé složky jejich těl. Zatímco většina biomasy lesa se skládá ze dřeva, které je nepoživatelné pro některé živočichy, mimo larvy xylofágních brouků a některých termitů, naproti tomu většina biomasy travnaté savany je tvořena trávami a jinými bylinnými rostlinami. Ta může být zkonzumována, v čerstvé nebo mrtvé formě, velkým množstvím bezobratlých živočichů a obratlovců. V savanových lesích a stromovitých (parkových) savanách je produkce biomasy průměrem těchto dvou extrémů. Na Pobřeží Slonoviny ve Banco forest je podíl listové hmoty v nadzemní části biomasy vytvářené stromy 1,6% a v Pasoh forest v západní Malajsii jen 1,18%. Zatímco trávy vytváří až 40% biomasy stojících těl, ve stromovité savaně v Fété Olé, a v některých částech Serengeti v Tanzánii nejsou prakticky žádné stromy, pouze trávy (F. Bourliére a M. Hedley, 1970).
Listí trav a jiných bylinných rostlin nacházejících se v savaně je navíc lépe poživatelné pro hmyz a obratlovce, než stejné rostliny v tropickém deštném lese. Stromy deštného lesa vykazují bohatou „chemickou obranu“proti býložravcům. Stromy syntetizují množství druhotných rostlinných sloučenin – alkaloidů a polyfenolů (Janzen,1974,1975) . Je známo, že stromy rostoucí na půdách málo bohatých na živiny musí vydávat více energie při jejich shánění. Proto stromy syntetizují více toxických sloučenin, které odpuzují býložravce a zabraňují tak ztrátám svých listů.
Pro podporu této myšlenky svědčí některé práce srovnávající dvě místa tropického lesa v Africe. Na místech s nízkou úrovní živin v půdě obsahovaly listy vysoký obsah fenolických sloučenin. Naopak trávy neobsahují žádné druhotné sloučeniny v porovnání s jinými rostlinnými skupinami (Lamprey a sp.,1974).
Listí mnoha savanových stromů a keřů je též požíráno některými pasoucími se savci. Dokonce některé z jejich semen (např. lusky Acacia tortilis v Africe) jsou aktivně vyhledávány jako potrava mnoha kopytníků, kteří přispívají k jejich šíření (Lamprey a sp.,1974).
Ačkoli je většina primární produkce bezprostředně dostupná pro konzumenty, část primární produkce v savaně může být přesto „uschována“ alespoň na krátkou dobu. Vytváří tak „energetickou reservu“ pro některé konzumenty během období sucha. Například semena jsou produkována v nadbytku i pro potřebu zajištění a udržení populace travin, klíčících každý rok. Živočichové živící se zrním, hlavně mravenci, ptáci a hlodavci, proto představují hlavní trofickou kategorii v tropické savaně. Suchá tráva, která má nízkou výživnou hodnotu pro přežvýkavce, je zkonzumována termity a dekompozitory, nebo zničena požárem.
To je v kontrastu se situací v tropickém deštném lese, kde vysoká teplota a vlhkost vedou k rychlému rozkladu organického materiálu.
2.2.5. Dynamika ekosystému savany
Ve srovnání s tropickými lesy mají savany relativně jednoduchou strukturu s méně druhy. Několik druhů je dominantních a tvoří většinu biomasy savan. Ekosystém savan je dynamická jednotka a poruchy v systému můžou přinést rychlou změnu v populacích dominantních druhů. Změna populace dominantního druhu může mít dalekosáhlý vliv na stav celého systému vytvořením řetězové reakce mezi jednotlivými komponenty. Poruchy mají charakter abiotický nebo biotický.
V severním Senegalu byl popsán proces, jak výkyvy srážek můžou mít dalekosáhlý a rychlý vliv na hlavní složky systému. Například Galat a Galat-Luong (1977) srovnávali roční rozdíly v úspěchu reprodukce a mortality kočkodana zeleného Cercopithecus aethopicus sabaeus, s rozdílným rozložením srážek ve sledovaných letech. Vliv srážek na populaci kočkodana je nepřímý. Srážky ovlivňují primární produkci potravních zdrojů kočkodana.
Naopak v severním Senegalu popsal Poupon (1979), jaký vliv mají výkyvy podnebí na stromy a následné složky systému. Během roku 1972 mělo sucho značný dopad na listnatou část (zpožděné olistění, omezení doby aktivní fáze), na kvetení (nekvetení, pozdější kvetení a omezené množství kvetoucích stromů) a na tvorbu plodů (úplná absence plodů a nebo jejich nízká produkce) stromů. Sucho způsobilo ústup dřevnaté vrstvy během období 1970 až 1979. Došlo ke změně mikroklimatu, zvětšila se půdní eroze, snížila se úrodnost půdy a produkce bylinné vrstvy (Poupon, 1979).
Dalším příkladem změn v savaně jsou biotické odchylky v potravním řetězci, například propuknutím epidemie dobytčího moru. První velká epidemie dobytčího moru ve východní Africe se odehrála na konci minulého století. První vlna se objevila v roce 1890 a do roku 1892 vymřelo 95% populace dobytka. V důsledku toho propukl u lidí hladomor a epidemické choroby, včetně neštovic. Nepřímo tak v důsledku dobytčího moru došlo k redukci divokých a domácích býložravců. Lvy přivedl k lidožravosti. Toto vše přimělo lidi opustit vesnické oblasti. Buš se rozrůstala, moucha tse-tse rozšířila svůj areál výskytu a spolu s ní následovala trypanosoma. Toto vše mělo za následek pokles lidské populace. V roce 1960 se podařilo odstranit dobytčí mor na planinách Serengeti. To odstartovalo sled událostí. Odstraněním dobytčího moru se zdvojnásobil počet jednoletých telat pakoně. Jejich úmrtnost klesla ze 75% na 50%. Tak došlo k nárůstu populace pakoňů z 250 000 na 500 000 kusů během 6 let v období roku 1961 až 1967. Pakoni jsou dominantními býložravci v této oblasti. Populace buvolů narostla za toto období přibližně z 30 000 na 50 000 kusů. Naopak populace zeber zůstala beze změny. Zebry nepodléhají dobytčímu moru, protože nepatří mezi přežvýkavce (Sinclair,1979).
Odstranění dobytčího moru způsobilo stupňovité změny v explozivním nárůstu dominantního spásače – pakoně. Tato změna způsobila velké změny i u jiných složek ekosystému. Hlavní vliv měla tato změna na vegetaci planin a lesnatých oblastí i na jiné populace býložravců a predátorů ve změně jejich početního poměru.
Pláně Serengeti poskytly dobrý důkaz o silných vzájemných vazbách mezi několika složkami ekosystému savan. Objasnily, že změny několika hlavních složek mohou mít dalekosáhlý vliv na jiné složky systému (obrázek 1).
2.2.6. Souhrnná charakterizace savany
Savana ukazuje značnou proměnlivost, variabilitu v prostoru a čase. Tato proměnlivost je jejím nejdůležitějším znakem. Může se vyskytovat ve formách přecházejících z tropického lesa až po otevřenou step v tropické zóně. Dominantní životní formou jsou zde početné druhy trav a šáchorovitých, které soutěží s dřevinami o využití půdní vlhkosti a živin v půdě. Podnebí je charakterizováno střídáním období sucha a dešťů, na které je vázán hlavní vegetační růst. Občas je savana postižena požáry, které jsou spolu s býložravci a živinami v půdě hlavními modifikačními faktory. Savany představují mladý biom v geologickém měřítku, v porovnání s biomem tropických lesů. Nabídly některým tropickým organismům nové příležitosti, které se úspěšně adaptovaly daným podmínkám životního prostředí. Jakékoli rychlé a inovační změny zatěžují prostředí savany.
Savany jsou důležitým místem ekologické proměnlivosti a mají klíčovou roli v evoluci biosféry a mnoha jejích druhů, ze kterých se skládá. Proto mnoho primatologů a antropologů spatřuje počátky našeho druhu v savanách, kde první prohominidé přešli ze stromů do otevřené savany.
 |
| Slon africký. Klasická představa savany jako otevřené pláně se solitérními stromy a velými savci.(N.P. Chobe, Botswana) |
¤ C4 – rostliny: světlomilné rostliny s pozměněnou fází fotosyntézy tzv. Calvinova cyklu. Zde se nazývá Hatch – Slackův cyklus.
¤¤ r-stratég: organismy produkující velké množství potomků, bez důsledné ochrany a výchovy do stavu dospělce.
Citace:
Jeník J. (1975): Život tropických savan I, Živa, 32, č. 1, Academia, Praha, s.2-5.
Jeník J. (1975): Život tropických savan II, Živa, 23, č. 2, Academia, Praha, s. 42-46
Conseil Scientifique pour l´Afrique, Yangambie (1956)
Dále:
G.F. de Oviedo y Valdes (1535); Shorter Oxford Dictionary (1944); Francouzský slovník slov z roku 1863-72 a Robert (1964-65); Grisebach (1872); Drude (1890); Shimper (1898); Beard (1953); Unesco (1979); Klíma (1970); Aubréville (1949, 1962); Adjanohoun (1963); Jung (1969); Gerakis a Tsangarakis (1970); Bernhard-Reversat (1977); Noy – Meir (1979); Lamotte (1978); Pandeya a Jain (1979); César a Menaut (1974); Poupon (1976,1979); Morel (1969); F. Bourliére a M. Hedley (1970); Janzen (1974,1975); Lamprey a sp. (1974); Galat a Galat-Luong (1977) převzato z: Bourliére F. (1992): Ekosystems of the world – Tropical savanna, Elsevier, Amsterdam, s. 1-17
3. Geografické rozšíření savan ve světě
Savany jsou rozšířeny na obou polokoulích v tropické zóně. Proto se lze setkat s označením „savany tropické“. Užití termínu „tropický“ vypovídá o zóně ležící mezi obratníkem Raka a Kozoroha (23,5°j.š. a 23,5°s.š.). Tyto astronomické hranice však nejsou živými organismy respektovány. Savana je ohraničena ve směru od rovníku zapojeným tropickým lesem. Na severu a jihu, mezi 20° až 30°, je její rozšíření ohraničeno polopouštěmi a pouštěmi.
Největší plochu zaujímají savany v Africe, kterou pokrývají přibližně ze 40% celkové plochy. Nejrozšířenější jsou v oblasti Sahelu, ve východní Africe a na jih od řeky Zambezi. Na Madagaskaru přirozeně pokrývají celou západní část ostrova, i když v důsledku činnosti člověka se jejich plocha rozšiřuje i do oblastí východní poloviny ostrova.
V Jižní Americe se rozkládají rozsáhlé savany v Kolumbii, Venezuele a v Brazílii. V této oblasti mají savany své zažité lokální názvy brazilské „campos“ a orinocké „llanos“. Savany se vyskytují také v nížinných oblastech střední Ameriky, jižním Mexiku a na ostrovech Velkých Antil.
Dříve savany pokrývaly i rozsáhlé oblasti poloostrovů Přední a Zadní Indie. Ty jsou dnes značně pozměněny činností člověka.
V Austrálii leží savany v severní části kontinentu. Velké plochy jsou také na ostrovech Malé Sundy a na Nové Guinei. Na ostrově Nová Guinea je nalezneme ve vyšších nadmořských výškách v centrální části ostrova a na jižním pobřeží.
V důsledku půdních faktorů se můžeme setkat s azonálním výskytem savanám podobných biomů i v perhumidní zóně.
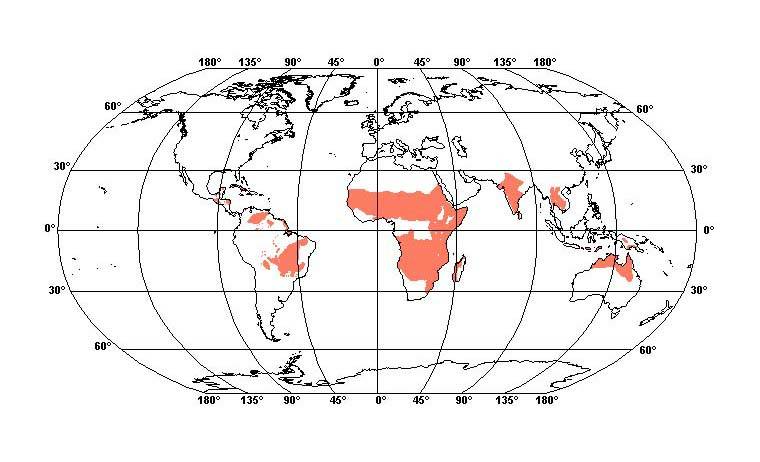 |
| Mapa rozšíření savana ve světě |
4. Africké savany
4.1. Fyzicko – geografické podmínky
Rozšíření savan na africkém kontinentu není náhodné, ale závisí na určitých činitelích: klimatických, edafických a biologických (viz. kapitola 4.3. až 4.5.).
Jednotlivé klimatické podmínky je těžké určit pro jejich vzájemnou souvislost. Sluneční světlo je nutné pro asimilaci. Teplota souvisí se světlem. Každý rostlinný druh má své teplotní maximum, optimum a minimum. I množství srážek určuje některé limitní charakteristiky pro růst rostlin.
Edafické podmínky jsou chemické a fyzikální vlastnosti půdy, která vytváří prostředí, ve kterém rostlina žije.
4.1.1. Klimatické podmínky
Hlavní výskyt africké savany odpovídá zóně, kde se střídá období dešťů a sucha, mezi vlhkou rovníkovou zónou a aridní zónou subtropů (mezi 10° a 30°j.š., resp. s.š.). Jejich hlavní výskyt leží v zóně mezi obratníkem Raka a Kozoroha (23,5°j. a 23,5°s.š.). Živé organismy ale nerespektují astronomicky definované hranice. Proto se v současnosti dává přednost teplotním hranicím, které lépe definují tropickou oblast. Tato hranice je určena nejnižší průměrnou teplotou nejchladnějšího měsíce 18°C. Dále se zde nesmí vyskytovat teploty pod bodem mrazu. Přesto ani teplotní hranice neposkytují přesné vymezení oblasti výskytu savan.
Podnebný systém tropických oblastí je důležitou částí globální atmosférické cirkulace. Hlavní hybnou silou tropické cirkulace je rozdíl teplot mezi rovníkovými oblastmi a subtropy. Schématicky byla tato cirkulace popsána jako Hadleyova buňka. Tento typ cirkulace se vyskytuje po celý rok pouze nad oceány. Hlavními složkami tropické cirkulace jsou: tropická zóna konvergence, pasáty, monzuny a tropické cyklóny. Převážná většina srážek spadne v období, kdy slunce je v nadhlavníku. Jejich množství závisí na vzdálenosti od tropické zóny konvergence a klesá směrem k tlakovým výším v subtropických šířkách.
Převládající proudění na severní polokouli má směr východoseverovýchodní, na jižní směr východojihovýchodní. Tyto větry se nazývají pasáty. Východní pobřeží Afriky ovlivňují též monzuny, které sem přináší od oceánu vydatné srážky.
Před tím, než se pokusíme přiřadit africké savany k některé klimatické klasifikaci, měli bychom si popsat hlavní klimatické prvky – sluneční záření, teplotu, výpar a srážky.
4.1.1.1. Sluneční záření
Sluneční záření je prakticky hlavním zdrojem energie pro planetární geosystém. Intenzitu přímého slunečního záření dopadajícího na horizontální plochu nazýváme insolace. Její hodnota závisí na zenitové vzdálenosti Slunce. Nejvyšší intenzity dosahuje v tropické zóně, kde její velikost je v suchých aridních oblastech, mezi 20°a 30°j. a s. š. Vysoký úhel slunce a bezoblačné nebe v těchto šířkách způsobuje, že zde průměrně dopadne 200 kcal cm-2rok-1 (@ 9GJ m-2rok-1). V důsledku vlhčího podnebí v rovníkových oblastech s množstvím aerosolů slábne množství dopadajícího slunečního záření. Intenzita slunečního záření je zde pouze 120 kcal cm-2rok-1 (@ 5GJ m-2rok-1). Savany jsou průměrem těchto dvou extrémů a typická úroveň intenzity slunečního záření je v Africe, na jih od rovníku 160 až 190 kcal cm-2rok-1 (@ 7-8GJ m-2rok-1) a na sever od rovníku 140 až 180 kcal cm-2rok-1 (@ 6-7,5GJ m-2rok-1) – H.A. Nix, 1992.
Šířková pozice savan určuje, že sezónní rozdíly v intenzitě slunečního záření v různých oblastech nemohou být větší. Největší rozdíly jsou vyvolávány střídáním období dešťů a sucha. Sezónní změny jsou vyvolány příchodem dešťů. Během období dešťů je intenzita slunečního záření v rozmezí 13 až 17 MJ m-2den-1 . Tato hodnota je stejná jako v oblastech vlhkých rovníkových pralesů. Během měsíců sucha dosahuje intenzita slunečního záření 20 až 25 MJ m-2den-1. Což je hodnota jakou denně obdrží aridní oblasti tropů okolo obratníků. Obecně africká savana obdrží největší množství slunečního záření koncem období sucha, a proto je zde v tuto dobu nejvyšší roční teplota.
Velká část africké savany se nachází v zóně, kde je velká intenzita slunečního záření. Amplituda intenzity záření je během roku nepatrná. Vrchol intenzity slunečního záření je na jaře, před tím než slunce dosáhne zenitu. Západní Afrika a rovníková východní Afrika má vrcholy dva, na jaře a na podzim, před sluncem v zenitu. V afrických savanách začíná období dešťů, když slunce dosáhne nadhlavníku. (H.A. Nix, 1992).
Měření předběžně vypočítaného slunečního záření poskytuje nezbytné podklady pro výpočty intenzity fotosyntézy , které jsou dnes používány pro modelaci růstu rostlin a přírůstku biomasy. Intenzity slunečního záření se též využívá k odhadu hodnot výparu.
4.1.1.2. Srážky
Hlavním znakem rozložení srážek v oblastech savan je jejich sezonalita. Hlavní vegetační růst souvisí se srážkami, naopak období sucha určuje ráz savanové vegetace. Savany se nejvíce vyskytují v zóně, kde roční srážky dosahují 1 000 až 1 500 mm (mapa 2). Reliéf, půda, podzemní voda a biotické vlivy mohou modifikovat makroklima tak, že ekosystém savany se vyskytuje i v přilehlých zónách, jinak příliš vlhkých nebo suchých.
Na okrajích tropické zóny, v chladnějších vrchovinách a ve vnitrozemí východního pobřeží, mohou být během roku srážky nižší. Tropická savana se zde vyvíjí i v místech kde úhrn srážek je okolo 500 mm nebo nižší. Na mělkých tvrdých podložních půdách nebo na extrémně propustných, se vyskytuje savana i za podmínek ročních srážek převyšujících 3000 mm. Představují tak oblasti savan s krátkým obdobím sucha. Extrémní varianta savan se vyskytuje na hrubých píscích s deficitem živin, a to v oblastech, kde není období sucha a jsou zde vydatné srážky.
Sezonalita rozšíření srážek je tedy hlavním a markantním znakem životního prostředí savan. Srovnávací index sezonality je koeficient variace (k.v. je standardní odchylka měsíčních srážek vyjadřující se jako procento měsíčních středních srážek). Hodnota nižší než 50 % vyjadřuje , že srážky jsou rovnoměrně rozloženy během roku. Hlavní výskyt v této kategorii je blízce shodný s rovníkovou zónou a naproti tomu i s mírnou vlhkou zónou vyšších šířek (H.A.Nix, 1992).
Koeficient odchylky (rozptyl variace) vyšší než 75 % značí silný sezónní kontrast srážek. Překvapující je, že velká část tropického pásma připadá do této kategorie. Do této zóny tak spadají prakticky všechny hlavní oblasti výskytu savan.
Index sezonality není postačujícím měřítkem během období dešťů nebo trvání období sucha. I místa s extrémně vysokými srážkami v rovníkových oblastech, jako je například jižní pobřeží Nové Británie na východě Nové Guinei, mohou mít vysoký index sezonality. Zde však jsou v průběhu celé sezóny srážky mnohem vyšší než výpar (H.A.Nix, 1992).
Sezónní rozložení srážek může poskytnout pohled na limitní množství vody, potřebné pro ekosystém savany. Mapa 3 ukazuje celkové srážky během nejvlhčích 6 měsíců, bez ohledu na aktuální čas. Hlavní výskyt ekosystému savan je koncentrován v místech, kde celkově srážky překračují 600 mm. Ty ale mohou být kompenzované různými faktory, jako je hlubší a více propustná půda, nižší výpar, hladina podzemní vody dosažitelná hlubšími kořeny stromů a keřů. V těchto oblastech mohou být srážky nižší a dosahovat pouze 400 mm. Horní hranice srážek není udávána. Regiony s více než 2000 mm během nejvlhčích měsíců mohou mít krátkou, ale intenzivní suchou sezónu. Ta se stává limitním faktorem pro rozvoj a souvislý výskyt stromů.
Mapa 4 ukazuje celkové množství srážek během nejsušších třech měsíců, bez ohledu na období výskytu. Poskytuje představu o podmínkách panujících během období sucha. V tropickém pásmu může výpar potenciálně dosáhnout během měsíců sucha až 200 mm. Ve třech měsících si to tak vyžádá 600 mm vláhy. Zbytek vláhy je uschován jako půdní voda, přibližně 150 mm. Žádné srážky během tohoto období nejsou vydatnější než 100 mm a vedou tak k nedostatku vody dosažitelné pro vegetaci (H.A.Nix, 1992).
Stále zelené lesy nerostou na místech, kde srážky během nejsuššího kvartálu nedosahují alespoň 100 mm. V ekosystému afrických savan jsou během nejsušších třech měsíců běžně srážky okolo 25 mm a někdy i 0mm.
4.1.1.3. Teplota
Vliv teploty na životní prostředí a tím na zonálnost vegetace byl zaznamenán již starými Řeky před 3 tisíci let. Hlavní globální klimatické klasifikace se zakládají na různých teplých, mírných a studených zónách. Výběr vhodných teplotních parametrů pro definování hranic představoval vždy problém pro vznik jednodušších klimatických klasifikací. Hranice byly často určeny přímým porovnáním izotermického gradientu s mapováním vegetačních pásem.
Ve většině prací je jako hranice hlavního rozšíření savan v Africe uváděna třicátá jižní a severní rovnoběžka. Savany se z hlediska klimatických klasifikací rozkládají převážně v tropické zóně.
Klimaticky definovaná tropická zóna vyniká vysokými teplotami vzduchu, jejich nízkým ročním kolísáním a skoro vůbec se zde nevyskytují teploty pod bodem mrazu.
Köppenova konvenční klasifikace například definuje tropické klima afrických savan na základě průměrné měsíční teploty vzduchu neklesající pod 18°C. Chybějí v něm chladná období. Roční amplituda teplot je zde malá (okolo 6°C). Budeme-li brát v úvahu pouze teplotní kritéria, může zde dojít k záměně s pásmem suchého klimatu, ve kterém dosahují průměrné teploty v několika měsících v roce18°C.
V nížinných oblastech Afriky jsou roční průměrné teploty (mapa 5) velmi vysoké, dosahují více než 24°C. Průměrné denní rozložení teplot všude v této zóně obvykle dosahuje průměrných ročních teplot. Tento nápadně široký zonální typ je narušen jen topografií terénu, prohlubněmi, blízkostí vody, přítomností teplých a studených mořských proudů, ostrůvkovitým rozložením vody a pevniny, relativní dominancí hlavních vzdušných mas a obecnou cirkulací atmosféry. Mnohé z těchto vlivů jsou patrné z mapy číslo 5, která znázorňuje průměrné roční teploty.
Na sever od rovníku je tento zonální typ teplotního gradientu narušen Etiopskou vysočinou a vrchovinami v západní Africe. Na jih od rovníku je teplotní gradient deformován severo-jižně se táhnoucími pohořími východní Afriky a angolskou vrchovinou. Vliv chladného oceánského proudu se uplatňuje na jihu Afriky, na jejím jihozápadním pobřeží (H.A.Nix, 1992).
Pro klimaticky definované tropické podnebí není proto vhodné používat hodnot průměrné roční teploty, ale spíše nejnižší průměrnou teplotu nejchladnějšího měsíce v roce (mapa 6). Tato hranice leží mezi izotermou 13°C až 18°C a souvisí s hlavním výskytem savan. Savany jsou přesto rozšířeny i v daleko chladnějších oblastech, kde průměrná teplota nejstudenějšího měsíce leží pod izotermou 8°C.
Na rozdíl od jiných vegetačních pásem není možné přesně vymezit teplotní hranice výskytu savan. Předpokládalo se, že tropická vegetace savan nesnese teploty pod bodem mrazu. Izoterma okolo 8°C je oblastí, kde se mrazy s určitou pravděpodobností vyskytují alespoň jednou ročně. Botanikové dnes soudí, že limitní není teplotní hranice, ale spíše délka období sucha.
4.1.1.4. Výpar
Voda se dostává do atmosféry převážně z aktivního povrchu (půdy, vodních ploch) fyzikálním procesem nazývaným evaporace. Výpar z rostlin se nazývá transpirace, která je fyziologickým procesem rostlinných orgánů. Oba pojmy jsou spojeny pod pojem evapotranspirace (celkový výpar). V přírodě nelze přesně změřit daný výpar žádným výparoměrem. Výpar ve výparoměru probíhá za umělých podmínek a může se výparu v přírodních podmínkách více či méně blížit. Přesto existují určité odhady výparu pro oblasti savan.
Během období dešťů je denní výpar mezi 2 až 4 mm, v období sucha denní výpar dosahuje 4 až 10mm. Celkový roční výpar je v oblasti savan za období dešťů a sucha 1 000mm, ale běžně dosahuje 1500 mm. Pouze v nejsušších okrajových částech savan je celkový roční výpar až 2000 mm (H.A.Nix, 1992).
     |
| Mapy jsou zpracovány dle The ekosystems of the world – Tropical savanna |
4.1.1.5. Klasifikace klimatu
Africké savany patří podle nejrozšířenější a nejuznávanější klasifikace W. Köppena do A – pásu vlhkého tropického klimatu. Tato teorie je založena na teplotním a srážkovém režimu a jeho vlivu na biotickou složku. V tomto pásu průměrné měsíční teploty vzduchu neklesají pod 18°C a roční amplituda denních teplot je malá (okolo 6°C). Roční úhrn srážek přesahuje 750 mm, srážky převažují nad výparem. Cirkulace atmosféry se vyjadřuje velkou pravidelností (pasáty a monzuny).
Vlhký tropický pás je podle této teorie dále členěn podle vztahu mezi ročním úhrnem srážek a úhrnem nejsuššího měsíce na Af – klima vlhkých tropických pralesů s rovnoměrným rozložením srážek během celého roku; Am – monzunový případ typu Af; Aw – podnebí savan s výrazně vyvinutým obdobím sucha, připadajícím na zimní období.
Přestože se africké savany vyvíjejí hlavně v podmínkách klimatického typu Aw, nalezneme je rozšířené v Am a mnohem méně vyvinuté i v Af, hlavně v oblastech, kde člověk narušuje přirozenou rovnováhu (H.A.Nix, 1992).
Jinou klimatickou klasifikací je genetická klasifikace B.P. Alisova. Tato klasifikace vychází z podmínek globální cirkulace atmosféry. Členění klimatu je dáno převládáním geografických typů vzduchových hmot. Klimatické typy jsou dále členěny na typy kontinentální, oceánské, podnebí západních a východních pobřeží pevnin.
Podle klasifikace Alisova patří africké savany do dvou různých klimatických typů. Za prvé do pásu rovníkových monzunů a za druhé do pásu tropického vzduchu.
Monzunové pásmo je charakterizováno vlhkým létem a suchou zimou s velkým rozdílem v množství srážek. Nejteplejším a nejsušším obdobím je jaro. Teploty vzduchu často přesahují 30 až 35°C. Hlavní minimum teplot připadá na zimu. Srážky vypadávají v podobě konvenčních lijáků a jejich množství klesá s rostoucí vzdáleností od rovníku. V závislosti na délce období dešťů klesá velikost srážek od 1000 – 1500 mm k 300 – 400 mm. V místech, například v Etiopské vysočině, dochází k jejich orografickému zesílení.
Tropický pás má typicky suché podnebí, s velmi horkým létem. Roční teploty dosahují 20°C a jejich denní amplituda je velká. Srážky zde vypadávají jen zřídka. Tento pás se rozkládá v okrajových oblastech savan v Africe. Patří sem severní okraje Sahelu a jižní okraje savan při polopouštních formacích Kalahari a středomořské vegetace typu Karroo.
Každá z těchto používaných klasifikací má své přednosti a nedostatky, protože se snaží zachytit celkové klima ve všech regionech světa a dochází tak ke značným generalizacím.
Rozšíření afrických savan je pravděpodobně určeno těmito klimatickými faktory:
1) roční celková intenzita slunečního záření dosahuje mezi 140 až 190 kcal cm-2rok-1 (@ 6-8GJ m-2rok-1)
2) roční průměrné srážky dosahují 1 000 až 1 500mm
3) sezonalita v rozložení srážek je vysoká, index sezonality je větší než 75 %
4) celkové srážky během nejvlhčích 6 měsíců dosahují 600 mm
5) celkové srážky během nejsušších 3 měsíců jsou menší než 50 mm
6) roční průměrná teplota vzduchu dosahuje 24°C
7) průměrná minimální teplota nejchladnějšího měsíce leží okolo izotermy 8°C
4.1.2. Geologické a pedologické podmínky
Matečnými horninami afrických savanových půd jsou horniny vyvřelé, metamorfované i usazené. Při vzniku půd se výrazně projevuje tropické zvětrávání a půdotvorné procesy. Svrchní horizont půdy má vlivem rychlého rozkladu živin nízký obsah humusu. Podle Půdní mapy světa FAO/UNESCO 1974 se na matečních horninách v tropických oblastech vyvíjejí hlavně nitosoly a ferasoly. Vedle jiných pedogenetických procesů v nich probíhá proces oželeznění, neboli feritizace a feralitizace. Dále zde ještě nalezneme Acrisoly, Luvisoly a Vertisoly.
K nitosolům patří fersialitické (železité) půdy (viz níže). Podmínkou pro jejich vznik je tropické nebo subtropické podnebí s dlouhým obdobím sucha, kdy půdy z větší části vysychají. Procesem oželeznění se uvolňuje velké množství Fe2O3, který často vytváří konkrece. Nejlépe vyvinuté železité půdy se vytvářejí v místech, kde srážky nepřesahují 1000mm a období sucha zde trvá až osm měsíců. V této půdní hmotě se nachází méně nezvětralých minerálů než ve ferasolech (feralitických půdách).
V suchých savanách se vyvíjí z železitých půd červenohnědé půdy a červenavě hnědé půdy. Značná část těchto půd tvoří přechod mezi půdami železitými a stepními. Průměrné množství srážek se pohybuje v rozmezí od 350 – 650 mm. Bylino – travní porost je řídký, stromy a keře nemají v době sucha olistění. V době dešťů dochází k procesu feritizace. Uvedené půdy lemují na jihu Saharu. Využívají se jako pastviny a při zavlažování a hnojení na nich lze pěstovat pšenici, sorgo, kukuřici i ovoce.
Druhou skupinou půd vyvíjejících se v podmínkách tropického klimatu jsou ferasoly. K této jednotce patří půdy vzniklé feralitizací – laterizací. Ferasoly vznikají ve stále vlhkém podnebí s přítomností rovníkových, tropických i subtropických lesů. Feralitické půdy mají velké množství volného Fe2O3 a Al2O3, které se zde hromadí následkem silného odnosu kyseliny křemičité. Nejtypičtější ferasoly nalezneme v rovníkových deštných lesích a ve vlhkých tropických lesích se srážkami nad 1200mm za rok. Méně typické feralitické půdy nalezneme ve vlhčích savanách.
Ferasoly jsou nejstarší a nejhlubší půdy na Zemi. Jsou velmi intenzivně zvětralé a jejich hloubka dosahuje 2 až 20 metrů, v některých případech i více. Svrchní vrstva půdy má nízký obsah humusu (okolo 1 až 3%), protože organický materiál je rychle rozložen a spotřebován bujnou vegetací.
V savanách nalezneme dva typy feralitických půd. Prvním typem je červená lateritová půda savan. Patřící většinou ferasolům i když část má charakter fersialitických půd. Vytváří se v horkém tropickém podnebí s výrazným obdobím dešťů a sucha. Roční úhrn srážek se pohybuje v rozpětí 1000 až 1500mm. Savany rostoucí v těchto klimatických podmínkách mají výšku travnaté vrstvy nad 1,5 metru. Určujícím pedogenetickým procesem je feralitizace, která probíhá během období dešťů. Během období sucha travnatý porost usychá a povrch se rozpaluje nad 50°C. Tím vznikají početné tvrdé železo-manganové konkrece velké 1 – 2 mm. Tak dostávají červené laterizované půdy „pseudopísčitý“ charakter. Množství rostlinného materiálu je značné, ale dochází k jeho rychlé mineralizaci. Proto je ve svrchní části půdy pouze 2 – 3% humusu. Červené lateritové půdy savan pokrývají rozsáhlé oblasti Afriky. Jsou poměrně úrodné, ale zatím jsou využívány primitivním zemědělstvím nebo vůbec ne. Při vhodném obdělávání a hnojení by poskytovaly vysokou úrodu zemědělských plodin.
Druhým typem feralitických savanových půd jsou laterity, nověji označované jako plinthin. Tento typ půdy má červenou nebo žlutavou barvu a silným vysoušením nápadně tvrdne (dehydratuje). Horizont s vysokým obsahem železa a hliníku může ztvrdnout v kompaktní slepenec. Vznikají tak snadno rozrušitelné půdní kůry a těžko rozrušitelné tvrdé pancíře. Jejich barva záleží na chemickém složení půdy. Vysoký obsah Fe obarvuje půdu do červena a fialova, světle červené až růžové půdy jsou bohaté na prvek hliníku. Přítomnost silikátů dokazuje zbarvení do šeda nebo béžova.
Vznik pancířů a kůry je dnes vysvětlován třemi způsoby. Obecně se soudí, že jde o souhru výhodné koncentrace a krystalizace sloučenin železa, která vede ke vzniku krystalické fáze oxidu železa. Kůry a pancíře jsou silné od několika centimetrů až po několik metrů. Často podléhají ničivým vlivům eroze. Pokrývají rozsáhlé plochy savan západní Afriky a Súdánu. Území pokrytá silnými kůrami a pancíři lateritu jsou prakticky ze zemědělského pohledu nevyužitelná.
Výskyt lateritického slepence v savanách způsobuje velkou proměnlivost životních podmínek pro různé rostliny a živočichy. Ovlivňuje mocnost půdy dosažitelnou pro rostliny jejich kořenovým systémem a také značně ovlivňuje půdně hydrologický režim. V místech, kde se vyskytuje vrstva lateritických slepenců, vznikají v době dešťů dočasná jezera nebo louže v depresích. Přechodně v nich žije vodní květena, například bublinatka. Po opadnutí vody vyrůstá na zbahnělém povrchu porost jednoletých trav (např. Loudetia kerstingii). V období sucha usychá vegetace na troud. Růst stromů je v těchto podmínkách nemožný. Toto vše ovlivňuje rozložení vegetace v savaně, místa s lateritickými slepenci jsou po většinu roku nápadné světliny uprostřed parkovitých a lesnatých savan. V západní Africe se tato místa nazývají „bowaly“(Jeník, 1975).
Acrisoly jsou kyselé silně zvětralé půdy převážně subtropického a též i tropického podnebí s horizontem argillic. Vedle jiných pedologických procesů v nich probíhá rubifikace spojená s feritizací.
Podle Půdní mapy světa FAO/UNESCO 1974 patří k půdám savan i luvisoly, které se však hlavně vyvíjejí v mírném humidním pásu v travnatých společenstvech. A podle tohoto znaku byly přiřazeny k půdám savan. Souvisí to však s nepřesností s definicí termínu „savana“.
Posledně zde jmenovaným, velmi častým typem půd jsou vertisoly, které vznikají na horninách bohatších na vápník, hořčík a jiné živiny. Vertisoly mají četné regionální názvy, například v Súdánu se nazývají badob. Jejich barva je tmavě černá. V hlubších vrstvách půdního horizontu se rozpadají na velmi jemné frakce a dávají tak prostor pro tvorbu druhotných půdních minerálů, například montmorillonitu nebo beidelitu. Tyto jílové minerály způsobují bobtnání a smršťování půdy a následné míšení jemnozemi podél vertikálních puklin v období sucha (vertické procesy). V Africe zaujímají velké plochy v severním pásu savan. Jsou to zpravidla těžké půdy a patří k nejúrodnějším půdám tropů.
Citace:
Jeník J. (1975): Život tropických savan I, Živa, 23, č.1, Academia, Praha, s. 2-5
Nix H.A. (1992): Ekosystems of the world – Tropical savanna, Elsevier, Amsterdam, 37 -61 s.
Půdní mapy světa FAO/UNESCO, 1974
 |
| Železité krusty s železomanganovými konkrecemi vytváří v půdách savany nepropustné podloží. Na druhé straně, kořenům rostlin zabraňuje v dosažení hladiny podzemní vody (Ngonye falls, Zambie) |
4.2. Typologie savan
Klasifikace savan se provádí na základě úplné floristické a fytocenologické syntézy. Analýza se provádí v druhé polovině období dešťů, kdy kvetou trávy a šáchorovité a kdy stromy i keře buď kvetou, nebo mají zralé plody. Z nedostatku informací se stále používá třídění na podkladě fyziognomickém, zejména podle poměrného zastoupení travin a dřevin (Jeník,1995).
V západní Africe Prof. Jeník zaznamenal, že ve směru od tropického lesa směrem k Sahaře se vystřídají tyto varianty savan: První jsou savanové lesy. Jsou to světlé háje s nižšími stromy a vysokostébelnými travinami v podrostu (savanové lesy jsou někdy označovány jako sezónní opadavé lesy nebo suché opadavé lesy; jsou to poslední stádia vývoje savan při okrajích deštného lesa). Ty jsou střídány stromovitou savanou, která má podobu parkovitého biomu se solitérními stromy a vysokými travami. Dále následují křovité savany s nízkými stromy a keři v souvislém porostu trav. Na ně navazují travnaté savany s převahou vysokostébelných trav a nakonec se zde rozprostírají „pseudostepi“. Jsou to nízkostébelné savany na hranicích polopouště.
Savany lze tedy rozdělit na:
· krátkostébelné savany
· dlouhostébelné savany
· křovité savany
· stromovité savany
· savanové lesy.
Naproti tomu, vegetační kategorie biomů afrických savan, vypracovaných koncilem v Yangambii, uvádí pouze fyziognomické čtyři typy (tab.1). Mapa 7 ukazuje výskyt jednotlivých typů savany na africkém kontinentu.
| Anglicky | Francouzky | české ekvivalenty |
| Savanna | Savane | Savana |
| Grass savanna | Savane herbeuse | Travnatá savana |
| Shrub savanna | Savane arbustive | Křovitá savana |
| Tree savanna | Savane arboreé | Stromovitá savana |
| Savanna woodland | Savane boiseé | Savanový les |
Tab.1 Vegetační kategorie afrických savan (podle koncilu v Yangambii)
Porovnáním těchto dvou, u nás a v anglofonním světě nejpoužívanějších fyziognomických klasifikací, obr. 4, není žádný rozpor. V typologii Prof. Jeníka je pouze navíc rozčleněna travnatá savana na další dva podtypy – dlouhostébelnou a krátkostébelnou savanu. I zde se však místy vyskytují solitérní stromy nebo keře.
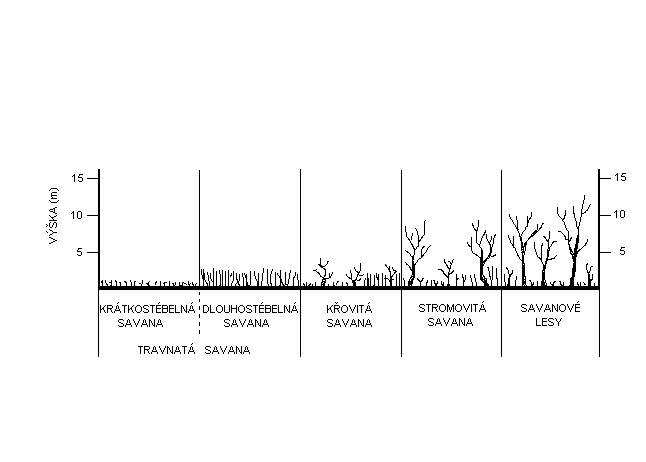 |
| Typy savan v Africe |
Existují i jiné typy klasifikací afrických savan, které se jen zanedbatelně liší od výše jmenovaných, a proto zde nebyly zmíněny.
Z hlediska zonálního zeměpisu klesá s ubývajícími srážkami podíl stromové složky. Proto by se savanové lesy a stromová savana měla rozkládat v blízkosti tropického deštného lesa a travnaté savany při okrajích polopouští.
Zonálnost jednotlivých typů africké savany je ale někdy výrazně narušena lokálními vlivy. Orientace svahů se v tropické zóně moc neprojevuje. Může se zde však značně projevovat orientace podle směru převládajících větrů. Příkladem je sahelská a guinejská oblast, kde svahy orientované k severu jsou bez dřevné složky, neboť v období sucha zde vane suchý vítr harmattan.
V oblasti úpatí svahů a údolních niv, kde se vyskytují lateritické půdy, je na místo lesních formací savana. Ve sníženinách bez odtoku vody se tvoří savany se solitérními palmami.
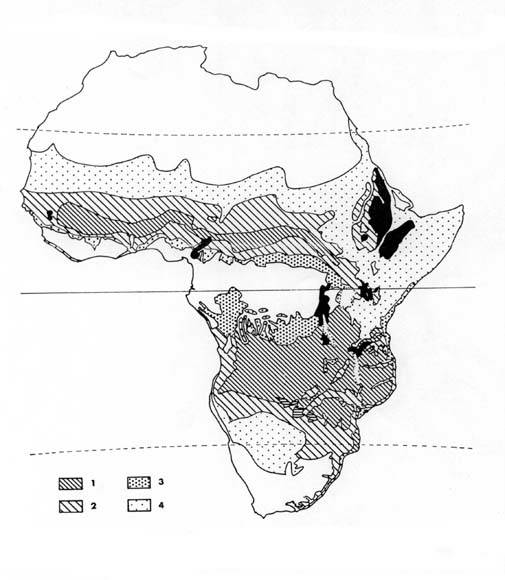 |
| Mapa hlavních vegetačních typů savany:1-tropické sezónní lesy (savanové lesy), 2-stromovitá a křovitá savana, 3-lesnaté formace a mozaika les a savana, 4-travnatá savana, černě horské oblasti (dle Ekosystems of the world- Tropical savanna) |
Citace: Jeník J. (1995): Ekosystémy (Úvod do organizace zonálních a azonálních biomů) – skripta, 1. vydání, Karolinum, Praha, s. 135
4.3. Vegetace africké savany
4.3.1. Stručná charakteristika
Přesně definovat vegetační kategorii savan a její rozšíření není pouze vědeckým zájmem. Je to též problém ekologický, protože spousta lidí popisuje tento vegetační typ jako „travnatou oblast“,“step“,“parkovou savanu“,“les“ atd., bez její následné definice. Mnoho klasifikací je založeno na rozličných kritériích (floristických, fyziognomických, klimatických a edafických), které mají rozdílnou váhu v různých vegetačních kategoriích.
Užití slova savana v širokém smyslu zahrnuje, na základě jednotného fyziognomického typu, lesnaté a travnaté oblasti tropického a subtropického pásu, včetně chudé krátkostébelné savany Sahelu, sezónních bažin dambos , savanového lesa miombo i suchých křovin.
Navzdory zřejmým rozdílům mají všechny tyto rostlinné formace společné ekologické rysy. Savany se odlišují od biomu tropického deštného lesa i od biomu polopouští a středozemního biomu množstvím klimatických, floristických, faunistických znaků a využitím půdy.
Někdy je však velmi složité určit v regionálním měřítku hranice jejích sousedních biomů. Tropický deštný prales někde zasahuje hluboko do biomu savany, často se podél tekoucích řek rozkládá les a občas zbytky lesních ploch vytváří společně se savanou mozaiku les – savana. Tyto dva biomy jsou někdy odděleny ostrými fyziognomickými a floristickými hranicemi. V sušších částech tropů a subtropů nalezneme v důsledku půdních podmínek mozaiku savana – poušť. Mezi těmito biomy je opět ostrá hranice. V důsledku každoročních klimatických změn se savana může změnit v poušť, v nízké křoviny (macchie) nebo suchou step. Vlastnosti půdy mění též rozložení savany.
Mapa 7 ukazuje rozšíření savan v Africe. Savana je na jihu a severu ohraničena pouštními a polopouštními formacemi. V blízkosti rovníku je pás tropického deštného lesa západní Afriky a konžského lesního bloku. V pásu savany se též vyskytují horské oblasti a mokřady.
4.3.2.1. Floristická příbuznost flory africké savany a fytogeografie
Zřejmá příbuznost je mezi suchomilnými rostlinami africké flóry s obdobnými v Asii a dokonce i Americe. Pantropické druhy jsou vzácné a jsou prezentovány mnoha druhy na všech třech kontinentech a ještě více čeleděmi. Tato situace není výsledkem pouze pasivního transportu a velikostí rozšíření jednotlivých druhů, ale hlavně velmi starého pantropického spojení, předcházejícího diferenciaci endemických skupin. Floristická příbuznost je hodně zřetelná mezi Afrikou a jižní Asií. Některé suchomilné druhy můžeme nalézt jak v nejzápadnější oblasti savany v Africe, tak i v oblasti severní Indie; to může být vysvětleno pouze dřívějším spojením obou kontinentů – v jižním kontinentu Gondwaně. Floristickou příbuznost mezi Afrikou a Amerikou můžeme vysledovat již ve třetihorách. Je nutno říci, že to neplatí pro druhy sem zavlečené člověkem, jako je např. Calotropis procera a Hyparrhenia rufa. Floristická příbuznost Afriky s Austrálií není tak zřejmá a početná – například Acacia spp., Adansonia spp. a Cochlospermum spp.(J.-C. Menaut, 1992).
 |
| Charakteristickým stromem africké savany je baobab. Je to majestátný strom, poskytující jedlé plody. Jako jeden z mála afrických stromů se dožívá vysokého věku (Nata, Botswana) |
Mnoho savanových rostlin s velkým rozšířením (zvláště ruderální a mokřadní druhy) jsou stromy a křoviny (Acacia spp., Cochlospermum spp. Jatropha spp. Prosopis spp. a Ziziphus spp.) – J.-C. Menaut, 1992. Týká se to též množství rostlinných terofytů*.
Z hlediska regionální fytogeografie náleží africké savany do paleotropické oblasti (Paleotropis). Rozsahem je to druhá největší, ale floristicky vůbec nejbohatší oblast. Má celkem 40 endemických čeledí. Z tropických rodů celého světa je 47% vázáno výhradně na její území.
Paleotropická oblast je rozdělena na tři části: africkou, indicko-malajskou a polynéskou.
Určitou podobnost je možné zjistiti i s neotropis. Některé rostliny typické pro tuto oblast se též vyskytují v Africe. Je to jen jeden druh bromélie Pitcairnia feliciana, který je striktně vázán na pohoří Fouta Djallon v Guinei a jeden epifitický nebo orofytický kaktus, Rhipsalis cassutha, rozšířený na africkém kontinentu. Netýká se to však ananasu Ananas comosus a Opuntia app., které sem byly zavlečeny člověkem a které zde zplanily. Opuncie se však v savaně nesnadno uchytává ( Chavalier, 1933).
Africká část je v bohatství květeny až na druhém místě paleotropické oblasti. V důsledku aridnějšího podnebí má Afrika relativně malé bohatství flóry (asi 30 tis. druhů). Veškerá vegetace Afriky se dělí na šest podoblastí: Africko-asijských pouští; Senegalsko-súdánskou; Východoafrických vysočin; Západoafrickou; Jihovýchodoafrickou a Jihoafrickou. K této oblasti patří i Madagaskar, který je rostlinami blízce příbuzný Malajské oblasti Paleotropis. Pouze podoblasti Senegalsko-súdánská, Východoafrické vysočiny a Jihovýchodoafrická jsou místem výskytu savan. Okrajově i Jihoafrická a Západoafrická podoblast. Zvláštní floristickou oblastí je Kapská oblast (Capensis) nenáležící k Africké části Paleotropis.
4.3.2.2. Geografické rozšíření flóry savany
Savana tvoří zřetelnou fytogeografickou jednotku, mimo bariéru tvořenou blokem tropického deštného lesa v západní a centrální časti, v sušších částech Afriky. Většina druhů savanové vegetace roste v celém pásu savan, od západu k východu a jihu. Jiné druhy mají bipolární rozšíření a nalezneme je pouze na severu a jihu, ale ne ve východní části Afriky. Nalezneme zde i jiné typy v rozšíření některých druhů. Ty mohou růst v pásu od Senegalu po Somálsko, jiné zase od Somálska k jihu.
Mnoho druhů savanových rostlin je rozšířeno okolo západního a středozápadního bloku deštného lesa. Patří do velkého množství rozdílných čeledí. Jsou to např. Actiniopteris radiata (Adiantacaea), Aristida hordeacea (Poaceae), Combretum aculeatum (Combretaceae), Grewia villosa (Tiliaceae), Parinari curatellifolia (Rosaceae), Piliostigma thonningii (Caesalpiniaceae), Acacia albida (Mimosaceae) a Diospyros mespiliformis (Ebenaceae). Jejich rozšíření okolo deštného lesa ve tvaru půlměsíce lze snadno vysvětlit historií geograficky kontinuálního rozšíření savany.
Výjimkou tvoří druhy nerovnoměrně rozšířené na sever a jih od deštného lesa a které se nevyskytují ve východní části Afriky. Tento typ rozšíření je zřejmě usnadněn šířkovou zonalitou biomu savany na severu a jihu od rovníku. Fyziografické a klimatické změny během čtvrtohor vysvětlují absenci některých rostlinných druhů v oblasti východní Afriky. Je zřejmé že východní koridor byl v minulosti několikrát přerušen a došlo zde k vymizení některých druhů, které jinde tvoří typickou vegetaci savan.
K podobným klimatickým změnám mohlo docházet i na západním pobřeží, kde mohl vzniknout koridor západ-jih, protože některé složky flóry Jihoafrické podoblasti (Kalahari – Namib) jsou příbuzné flóře západní Afriky.
Floristická podobnost severní a jižní části savany je značná 63% rostlinných rodů rostoucích v zóně Sahelu koresponduje s oblastí jižní Afriky (Monod,1971). Též zde nalezneme podobnost , floristickou a fyziognomickou, mezi savanovými lesy na jihu (mezi Isoberlina, Monotes a Uapaca) a severu (Brachystegia, Isoberlina a Jubernardia – J.-C. Menaut, 1992).
4.3.2.3. Životní adaptace flóry savany
Rostliny tropické savany Afriky se liší od flóry deštného lesa množstvím morfologických a funkčních charakteristik, které nejsou pouze podmínkami fyziognomie vegetace, ale mají též vlastní vývoj a produkci.
Fenologie
Savanové stromy se skládají převážně z opadavých druhů. Opad listů v období sucha dává lesnatým společenstvům charakteristický sezónní vzhled. Opadávání listí je bezpochyby přizpůsobení rostlin na sucho a jeho trvání záleží na vodních podmínkách prostředí. Některé druhy proto mají na vlhčích pláních listí déle než na sušších svazích.
Většina stále zelených formací v biomu savany se vyskytuje pouze v místech, kde je během období sucha dostatečné množství vláhy. Některé stále zelené keře přežívají i na suchých stanovištích, kde půda má nízkou schopnost vytvářet konkrece (neprobíhá zde oželeznění ani laterizace).
Stromy mají dvě strategie, jak přežít období sucha: první z nich mají xeromorfické listy a druhé mají listy mesomorfické, opadávající během období sucha. V savaně jsou díky svým lepších kompetivním schopnostem dominantní druhy opadavé. Ačkoli jejich listy jsou produktivní pouze krátký čas, je pro jejich vznik požadováno malé množství materiálu. Opadavé druhy rostou mnohem rychleji, jsou lepší producenti a mnohem úspěšnější v kompetitivních situacích.
Běžným znakem savany je otevírání pupenů bylinných i stromovitých druhů na konci nebo dokonce na vrcholu období sucha. V oblastech velmi sezónního klimatu může být opad listů velmi nepravidelný. Extrémem je Acacia albida, která se pokrývá čerstvými listy v období sucha a ztrácí je s příchodem doby dešťů.
Sexuální rozmnožování
Je obecně přijímáno, že světlomilné rostliny mají mnohem větší a více jasnější květy, než druhy stínomilné. Náhlé rozvití je častým jevem stromů savany (Leguminosae), křovin (např. Cochlospermum) a bylin (např. Cuculigo). Není to však pravidlem. Mnoho stromů savany má malé a diskrétní květy. S malou výjimkou v jižní části kontinentu.
Bylinné rostliny kvetoucí ve společenství stromů rozkvétají během období sucha, protože jsou stromy bez listí. Jejich květy jsou tak více nápadné. Mnoho z nich též kvete počátkem období dešťů.
Spouštěcí mechanizmy kvetení mají mnoho možných synchronizací, změny ve fotoperiodě, okolní teplotě, požáry a živiny v půdě, ale žádný z nich sám o sobě nemůže být považován za spouštěcí mechanizmus kvetení všech savanových druhů. Kvetení po požárech u některých geofytů**** zde není bráno v úvahu. Například štětkovka trojmužná (Themeda triandra) klíčí výhradně po požárech savany. Před vyrašením se její semena působením teplotních změn aktivně zavrtávají do půdy. Přesto velmi málo savanových bylin kvete během období sucha. Většina druhů vykvétá až během období dešťů.
Některé druhy rostlin kvetou stále v ten samý čas. Například v savaně Guinei na Pobřeží Slonoviny vykvétají všechny hemikryptofyty** na konci svého vegetativního cyklu, tj. konec období dešťů (J.-C. Menaut, 1992).
Rozšiřování semen větrem je v otevřeném prostředí savany velmi důležité. Semen takto adaptovaných (mají křidélka) je v savaně velké množství (např. Combretaceae). Semena u některých rodů rostoucích v lesích jsou mnohem menší než u jejich savanových protějšků.
Rozšiřování semen externím transportem živočichy ( v srsti, na kůži) je velmi rozšířeno v oblasti savan Sahelu, kde mnoho semen má háčky nebo přilnavé vlásky (např. Cenchrus biflorus, Guiera senegalensis – J.-C. Menaut, 1992).
Vegetativní rozmnožování
Výskyt stromovitých rostlin rozšiřujících se z plazících se podzemních výhonků je též častý. Nadzemní časti mají vytvořené z polodřevních stonků. Tyto rostliny vytváří v nadzemní části husté shluky, podobné travním trsům nebo svazkům. Po poškození výhonky raší z kořenových prutů a mohou pokrýt plochy o několika metrech čtverečních.
Někdy jsou rodičovské rostliny stále spojeny s ostatními. To je případ Piliostigma thonningii. Vedlejší kořeny se rozšiřují pod zemí do okolí. Z nich vyrůstají adventivní kořeny a následně mladé rostliny, které jsou zdánlivě samostatnými rostlinami (J.-C. Menaut, 1992).
Pravé vegetativní rozmnožování nalezneme u Landolphia tholloni, Ochna arenaria a Parinari pumila (Makany,1976). Jejich podzemní části vysílají do okolí adventivní kořeny, které se na různých místech rozkládají a dávají vyrůst novým individuálním rostlinám.
Vegetativní rozmnožování nehraje nezbytnou úlohu ve znásobení stromů v savaně.
4.3.2.4. Morfologie savanových druhů rostlin
Nadzemní část
Životní strategie stromů jsou v savanách odlišné od druhů tropického lesa. Žádné patro jejich vývoje korun a kmenů nikdy nedosahuje velkých výšek. Mají nízké větve a mohou se větvit hned u základu. Koruny jsou velmi široké a mnohem více vyvinuté než kmeny. Životní zvyky jsou velmi rozličné, ale individuální tvary lesních druhů společně s křovinami a způsob větvení jsou podobné. Keře někdy dosahují stejných výšek jako stromy. Tvary mohou být rozdílné i u stejného druhu stromovité rostliny. Některé stromy jako je Parkia bicolor, Parinari excelsa, Ceiba pentandra, které obvykle nalezneme v ekosystému lesů, jsou v prostředí savany zakrnělé, s nízko položenými větvemi. Typické stromy savany, jako je baobab Adansonoa digitata, mají obvykle základ kmenu široký a krátký. Na suchých stanovištích však mohou dosahovat až 20 m výšky.
Může se stát, že jednotlivé druhy se vyskytují v rozdílných růstových a životních formách, i když rostou v podobných ekologických podmínkách. Příkladem je Nauclea latifolia, která může vypadat jako plazivý keř nebo strom a Piliostigma thonningii, vyskytující se ve formě jako fanerofyt*** vyšší než 6 m nebo jako hemikryptofyt s obrovským kořenovým systémem.
Silné kůry většiny stromů slouží jako ochrana před častými požáry savany, např. Butyrospermum paradoxum a Cussonia barteri (J.-C. Menaut, 1992). Přesto i mnoho druhů stromů se slabou kůrou roste v savaně úspěšně.
U savanových keřů a stromů je častý výskyt trnů a ostnů (např. Acacia spp., Balanites agyptiaca, Euphorbia spp.), hlavně ve křovinatých savanách v Sahelu. Poskytují ochranu před spásáním velkými savci. Přesto neochrání listy před spásači jako je žirafa (Giraffa camelopardalis) a nosorožec (Dinoceros bicornis). Je to též jedna z cest, jak zamezit ztrátám vody prostřednictvím evaporace. Přesto jejich výskyt, ze všech zastoupených stromů a keřů v suchých savanách, nedosahuje více než 10 % (J.-C. Menaut, 1992).
Listy stromů savany dosahují malých až středních velikostí. Mikrofilní druhy jsou častěji zastoupeny v suchých klimatických oblastech. Rostliny s velkými listy nejsou v afrických savanách zastoupeny vůbec, kromě vějířovitých palem (Borassum aethiopum a Hyphaene thebaica). V tomto prostředí lze často nalézt tvrdé listy se silnou vrstvou kutikuly. Většina druhů má listy měkké, dokonce i v aridních podmínkách. To je shazováno v období sucha. Savanová flóra je většinou tvořena druhy, které ztrácí své zelené hydrofilní části, zatímco více xerická flóra pouští a mediteránních oblastí má větší procento rostlin s pravými xeromorfními trvalými orgány (Schimper,1903).
V období, kdy stromy shazují své listy, shazují též často své roční výhonky, což je důvod jejich pomalého růstu.
Xeromorfní adaptace rostlin savany představuje množství morfologických a fyziologických adaptací, které zamezují ztrátám vody (silná epidermis, zanořené a chráněné stoma, malé listy atd.). Dále sem patří také zvýšené uchovávání vody (kaktusům podobná Euphorbia spp., vodu uchovávající silné kmeny baobabu Adansonia digitata nebo Adenium obesum). Rostliny vykazují v adaptaci na životní prostředí množství rozličných strategií.
Životní a růstové formy známé z tropického deštného lesa se vyskytují v africké savaně zřídka. Příkladem jsou jen parazitické rostliny. Epifyty jsou velmi vzácné společně s několika druhy kapradin, orchidejí a játrovek. Liánovité druhy jsou početnější : úzké stromové liány nalezneme dokonce v suchém savanovém lese. Plazivé druhy a vína (např. Lochocarpus cyanescens, Acacia pennata, Strophantus sarmentosus) se zde vyskytují často. Xeromorfické a často sukulentní liány s velmi malými listy nebo bez listí (např. Cissus quadrangularis, Seyrigia gracilis) se často vyskytují i v sušších oblastech. Běžné jsou zde též popínavé rostliny (např. Cucurbitaceae – J.-C. Menaut, 1992).
Podzemní část
Oproti deštnému lesu mají stromy v savaně dobře vyvinutý kořenový systém prorůstající do velkých hloubek.
Bylinné rostliny, hlavně jednoleté, mají také rozsáhlý kořenový systém, často vytvářející koberce v blízkosti povrchu. Až 80% kořenů je lokalizováno v hloubce do 30 cm pod povrchem (J.-C. Menaut, 1992). Pro uchování vody využitelné během nejsušších částí roku je potřeba využit co největší plochu půdy, aby kořeny zajistily potřebnou dodávku vody.
Dobře vyvinutý kořenový systém rostlinám dovoluje přežít sucho i požáry. Na konci vegetačního cyklu, před tím než začne období sucha, jsou voda a živiny uchovány. Nadzemní části rostlin zvadnou. Kořenový systém dovoluje, jako velký rezervoár, rychlý růst vzrostlých vrcholů. Před tím než začne fotosyntéza se rychle prodlužují. Přesunem živin z kořenů do výhonků dosahuje kořenový systém nižších hodnot přírůstku biomasy. Před příchodem dešťů nejsou absorpční kořínky aktivní. Kořenový systém rostlin savany je velmi citlivý na vodní podmínky v půdě. Růst biomasy kořenů může být snížen, kdykoli během roku, když se vyskytne sucho. Podzemní orgány reagují na sucho stejným způsobem jako nadzemní části, ovadnutím jejich hodně jemných částí. Tato reakce může být velmi rychlá.
Nízké stromové rostliny reagují stejně jako jednoleté rostliny. Na rozdíl od nadzemní části, která je buď jednoletá nebo částečně jednoletá, je nadměrně vyvinut jejich kořenový systém. Mladé semenáčky mají rychle rostoucí kořenový systém, který zvyšuje jejich šanci přežít. Roční vadnutí nadzemních orgánů vyčerpá rostliny, které nemají dostatečný kořenový systém. Dobrý kořenový systém dovoluje po opadnutí listů rychlé rašení (po suchu, ohni, spasení nebo poškození). Některé druhy přežívají jako hemikryptofyty s dobře vyvinutým kořenovým systémem. U většiny fanerofytů je poměr kořeny/výhonky ve prospěch prvního z nich. Tento poměr se s věkem postupně snižuje. Dokonce i u stromů a keřů v Sahelské zóně (J.-C. Menaut, 1992).
Většina stromů a keřů savany má kořeny rostoucí do značných hloubek, od kterých se oddělují postraní kořeny. Růstem do hloubky se snaží dosáhnout hladiny podzemní vody, což jim usnadňuje přežití. Není to však pravidlem, protože rostliny rostoucí v aridních oblastech, mají kořenový systém spíše rozprostřen do plochy při povrchu půdy, aby absorbovaly vodu, která během slabých dešťů nasytí pouze svrchní část půdy. Toto je případ i většiny ostatních rostlin savany. Mají velmi mělký a horizontálně uložený kořenový systém a v případě, že mají hlavní kořen, ten nezasahuje do hlubších vrstev půdy. V písčitých savanách jižní Afriky, má kořenová biomasa v půdě podobné rozložení, ale hlavní kořen některých druhů stromů prorůstá do hlubších vrstev. Postraní kořeny obvykle převládají nad hlavním kořenem, zvláště u dospívajících stromů. Síť kořenů zde prorůstá do hloubky 10 až 20 cm a vytváří se tak labyrint kořenů stromů, křovin a trav.
Koexistence stromových rostlin s bylinnými je způsobena různým využitím půdní složky ekosystému. Stromy a keře využívají hlubší vrstvy půdy. Trávy, ostřice a jiné byliny využívají naopak mělčí vrstvy půdy.
4.3.2.5. Životní formy
Typickým znakem druhů rostlin v africké savaně je variabilnost životních forem. Podmínky životního prostředí se zde mění, proto některé druhy, např. Cissus, který byl určen jako liánový fanerofyt v lesním prostředí, se v savaně vyskytuje jako geofyt****(Tchoumé,1966).
To samé platí pro mnoho stromových rostlin Senegalo-súdánské a Jihovýchodoafrické floristické podoblasti (J.-C. Menaut, 1992). Odstranění jistého ekologického tlaku, jako jsou požáry a spásání, dovoluje některým druhům dosáhnout své původní životní formy.
V normálních podmínkách savany se rozdílné životní formy kříží každý s každým, a proto je zde klasifikace obtížná. Některý druh může být přiřazen jedním autorem jako hemikryptofyt, dalším jako víceletý terofyt a jiným jako chamaefyt nebo geofyt (J.-C. Menaut, 1992).
„Biologické spektrum“ životních forem představuje procentuální zastoupení jednotlivých druhů a jejich životních forem v prostředí savany. Tento ukazatel může mít i svou nespolehlivost, protože je užíván například pro celkovou fyziognomii savanových rostlin nebo pro určité regionální ekologické podmínky.
Spektra náležející k specifickým místům patřících jednotlivým regionům jsou někdy značně odlišná od situace v makroregionu. Jednotlivá místa mohou mít odlišná půdní složení.
Tabulka ukazuje srovnání „biologických spekter“ rozličných typů africké savany.
Množství fanerofytů se s ubývajícími srážkami moc nemění, ačkoli se lehce snižuje v suchých savanách. Pravděpodobně je to způsobeno regulační složkou ohně. Vlhké savany jsou zvláště bohaté na geofyty a hemikryptofyty; tyto dvě složky mají dobrou ochranu proti intenzivním ohňům vznikajícím ve vzrostlých porostech trav. Chamaefyty*****dobře rostou v savanách ležících v zóně mezi vlhkou a suchou savanou. Terofyty jsou nejpočetnější v aridních savanách.
V různých podmínkách rostou jak rostliny víceleté (stromy a keře) tak i jednoleté (zvláště trávy). Kooperace mezi jedno a víceletými, tedy bylinnými a stromovými rostlinami, je velmi důležitým znakem savany.
 |
| Akácie se vyskytují v křovité či stromovité formě. Akácie na obrázku je velkého vzrůstu a ověšena spoustou hnízd snovačů. (Okavango delta, Botswana) |
4.3.2.6. Kooperace mezi bylinnými a stromovými rostlinami
Ve většině biomů jsou stromy a bylinné rostliny antagonisté, ale savany jsou charakteristické výskytem obou forem.
Bylinné rostliny mohou existovat ve formacích stromů, ale i bez nich, to samé platí pro stromy. Přesto jsou stromovité rostliny mnohem citlivější k podmínkám životního prostředí, než např. trávy. V důsledku příhodných změn prostředí, pro jednu nebo druhou složku, se savana rychle mění v lesnatou nebo travnatou. Savana je tak vlastně „bojovou zónou“ mezi stromy a travami, a jejich proporce závisí na dostupnosti vody obsažené v půdě. Úspěšnost závisí na strategii hospodaření s vodou a kořenovém systému.
| Oblast (od shora dolů klesá množství srážek) | fanerofyty | chamaefyty | hemikryptofyty | geofyty | therofyty |
| Guinejská savana (vlhké klima) | |||||
| Pobřeží Slonoviny | 28 |   | 42 | 6 | 24 |
| Kongo | 14 | 25 | 26 | 16 | 19 |
| Nigérie | 30 |   | 23 | 21 | 25 |
| Sudáno-zambezijská oblast (přechodná savana) | |||||
| Středoafrická republika | 13 | 24 | 8 | 11 | 40 |
| Rwanda | 29 | 30 | 12 | 7 | 20 |
| Kongo (Zaire) | 38 | 44 | 9 | 4 | 5 |
| Sahelská savana (suchá savana) | |||||
| Mauritánie | 24 | 7 | 6 | 2 | 61 |
| severní Senegal | 19 | 2 | 2 | 2 | 75 |
| severní Čad | 14 | 2 | 12 | 5 | 67 |
Zpracováno podle Tropical Savannas, Ekosystems of The World, 1992
Tab.2 Procentuální zastoupení různých životních forem rostlinných druhů v některých afrických botanických oblastech
Listy trav uvadají, a tak v období sucha nepotřebují vodu k transpiraci. Spící rostliny též nepotřebují vodu a živé si ji musí shromáždit do podzemních částí. Trávy jsou dobře přizpůsobeny období sucha a nízkým srážkám, protože vodu potřebují pouze po vegetačním období. Voda je vstřebávána hustým kořenovým systémem těsně pod povrchem půdy.
Na druhé straně stromovité rostliny požadují vyšší a více pravidelné srážky. Dokonce potřebují vstřebávat vodu i během období sucha, aby vyrovnaly ztráty vody v důsledku transpirace svých nadzemních částí. Stromy proto mají velmi rozsáhlý a do značné hloubky zasahující kořenový systém.
Walter (1971) se pokusil na tomto základě vysvětlit relativní rozšíření bylinných a stromovitých rostlin v savaně. Na kamenitých půdách, které udrží jen malé množství vody a kde voda není jednotně rozložena a prosakuje do značných hloubek, převládají stromy nad travami. V půdách, které mají vhodnou strukturu pro absorpci a uchování vody ze srážek, dochází ke kompetiční rovnováze mezi stromy a srážkami a její ustanovení záleží na množství srážek. Když jsou srážky nižší než 250 mm za rok, stromy mizí a vyskytují se zde pouze chudé trávy bez stromové složky. V oblastech s režimem srážek mezi 250 až 500 mm za rok stále dominují trávy s občas se vyskytujícími stromy. Nad 500 mm srážek za rok je stále více stromů. Jejich podíl roste s množstvím srážek.Teprve když stromy vytvoří zápoj korun a stín zamezuje růstu trav, je narušena rovnováha. Stromové druhy pak značně dominují. Jejich výskyt záleží nejen na množství srážek a střídání období dešťů a sucha, ale i na půdních podmínkách. Stromy a keře nesnáší dlouhodobé zamokření půdy, zamezuje to transpiraci kořenů. V savanách delty Okavanga rostou stromy jen na termitištích vyvýšených nad okolní zamokřovanou půdu. Stromy rostoucí podél řek zde nachází dobré podmínky, protože půda je zde zamokřená jen krátkou dobu. V některých savanách Afriky pouze palmy (Borassus aethiopum, Hyphaene thebaica, Phoenix reclinata) a některé druhy akácií odolávají záplavám.
Chudé půdy mají vliv i na výskyt savany v tropickém lese, jako jsou savany esobe v pánvi řeky Kongo (J.-C. Menaut, 1992). V oblasti Guinejského zálivu se též vyskytuje savana v podmínkách, kde by měl být tropický deštný les. Velmi chudé laterické půdy nedovolují opětovné zalesnění.
Silný tlak člověka a jeho stád, které nadměrně spásají vegetaci, dále také požáry, nedovolují růst stromové vegetaci. Též trávy Andgropogoneae a Paniceae (Pennisetum purpureum) nedovolují růst stromů, protože jsou při občasných požárech dobře hořlavé (Jacques-Félix, 1959).
V dnešní době je v oblastech Sahelu západní Afriky nízký výskyt stromů, protože během velkých such v této oblasti v nedávné minulosti došlo k velké mortalitě stromů. Sucho nedovolovalo nahrazovat ztráty způsobené člověkem, který potřeboval k otopu palivové dříví.
4.4. Hlavní vegetační kategorie savany
4.4.1. Uzavřené formace
Tímto termínem je myšlen zápoj korun stromů a keřů, které zakrývají nebo znemožňují větší rozvoj travního pokryvu.
Suché opadavé lesy
Někdy jsou označovány jako tropické sezónní lesy. Jejich fyziognomie je podobná deštnému lesu. Ztrácí listí na několik týdnů v roce. Podrost je většinou také opadavý. Tyto tropické lesy rostou v suchých oblastech bez větších klimatických extrémů. Jejich druhy nemají nic společného s druhy obývajícími vlhčí lesy. Bez ohledu na jejich sporý travní pokryv se v nich často vyskytují požáry,. Jejich degradací se z nich stává stromovitá savana nebo otevřené savanové lesy. Tvoří poslední stádium vývoje savan směrem k deštnému lesu. Proto jsou přiřazeny k savanovým lesům. V dnešní době se vyskytují jen jako relikty, protože je na ně značný tlak požárů a těžby.
Suché opadavé lesy jako biom savany nesmí být směšovány s vlhkými poloopadavými lesy při okrajích deštného lesa, které jsou občas také nazývány západoafrickými autory „suché lesy“- anglický překlad (J.-C. Menaut, 1992).
Stálezelená facie
Některé stálezelené rostliny rostou i mimo suché opadavé lesy. Vytváří stálezelené lesy, které se též nazývají „husté suché lesy“ a vyskytují se v okolí Guinejského zálivu. Snadno přechází ze svého klimaxového stádia ve stromovou savanu. Stálezelené lesy v jižním Kongu (Zairu) nejsou příliš vázány na typ půdy. Množství srážek musí být větší než 1300 mm během 4 měsíců jdoucích po sobě. Tyto lesy jsou bohaté na druhy Entandophrygma develoy a Diospyros holeyma a v okolí je množství jiných mechů a játrovek (J.-C. Menaut, 1992).
Na jihu střední Afriky se suché stálezelené lesy vyskytují na půdách poskytujících v době sucha (trvá 7 měsíců) dostatečné množství vláhy. Tyto lesy jsou tvořeny třemi patry vegetace. Nejvyšší tvoří stromy se zápojem korun vysokých až 25 m. Většina rostlin druhého patra je vysokých 10 až 15 m. Pak následuje třetí patro tvořené 5 m vysokými keři. Šplhavé druhy rostou přes všechna tři patra. Povrch je holý nebo pokrytý trávami deštného lesa. Byly popsány tři typy tohoto lesa. V prvním dominují druhy Parinari- Syzygium, ve druhém Cryptosepalum pseudotaxus (severní část Kalahari na písčitých půdách) a ve třetím Upaca spp.
Stálezelené lesy v súdánské zóně se vyskytují dnes jen na skalnatých místech bez výskytu požárů. Hlavní patro roste do výše 15 až 25 m a je tvořeno jen jedním druhem Gilletiodendron glandulosum nebo Guibourtia copallifera. Povrch je pokryt trávami.
V místech, kde bylinnou vrstvu tvoří savanové trávy, se vyskytují také lesy, tvořené Uapaca togensis. Též v suchých a stromovitých savanách Zambie převládají stromy rodu Uapaca spp.
 |
| Stálezelená složka savanového lesa tvořená pryšcovitými (Euphorbia sp.) rostlinami (N.P. Kruger, Jižní Afrika) |
Opadavá facie
Opadavé lesy jsou považovány za regresivní stádium stálezelených lesů. V západní části kontinentu se vyskytují jen ostrůvkovitě. Jsou tvořeny hlavně savanovými druhy s množství světlomilných trav (např. lesy Albizia-Anogeissus nebo Parkia-Pterocarpus). Ve východní části Afriky se vyskytují lesy tvořené stromy Isoberlinia a Anogeissus, s bylinnou vrstvou.
Při dobrých podmínkách (větší množství srážek, nevyskytování požárů) snadno přechází otevřená savana v uzavřené formace suchého opadavého lesa a naopak. V jižní části střední Afriky se vyskytují lesy tvořené druhem Baikiaea plurijuga, které rostou v horkém a suchém podnebí. Srážky zde dosahují pouze 650 mm, ale hluboká písčitá půda má dobré retenční schopnosti (J.-C. Menaut, 1992). Vyskytuje se však pouze v místech s malým množstvím požárů. Ve spodních patrech roste Combretum, Acacia a Commiphora spp. Rozkládají se většinou mezi opadavými lesy s dominantním rodem Brachystegia a křovitou savanou tvořenou rodem Acacia.
Husté keře
Křoviny na některých místech africké savany vytváří formace podobné suchým opadavým lesům s hustým zápojem korun. Travnatá vrstva je zde nesouvislá. Rostou převážně na místech chráněných před účinky požárů savan. Často se vyskytují na pobřeží (např. Chrysobalanus v Ghaně nebo Strychnos v Angole). V západní Africe, kde je vlhčí podnebí, je složkou těchto hustých křovin i bambus (Oxytenanthera abyssinica).
V jižních částech střední Afriky vytváří tyto husté keře pásy obklopující suché stálezelené lesy.
Ve východní Africe v Tanzánii pokrývají keře typu Itigi velké plochy. Jsou tvořeny druhy Baphia, Bussea a Pseudoprosopis. Dosahují výšky až 5 m. Vyskytují se ve stejných klimatických podmínkách jako opadavé lesy, ale půda je zde více kyselá. V pobřežních oblastech severní části Viktoriina jezera jsou husté křoviny tvořené pryšci Euphorbia candelabrum.
Na jílovitých půdách středního Súdánu se vyskytují husté keře tvořené Acacia mellifera. V sahelské zóně jsou časté ostrůvkovité výskyty hustých křovin s druhy Acacia spp. od Somálska po Senegal.
Na štěrkových nebo lateritických půdách se vyskytují neproniknutelné „buše“ s druhy Pterocarpus lucens a Combretum spp.
Na chudých kamenitých a vlhčích půdách v pobřežních depresích na jihu súdánské oblasti rostou husté keře s Acacia ataxacantha smíšené s Combretum micranthum.
4.4.2. Otevřené formace
Jsou charakteristické výskytem dobře vyvinuté bylinné vrstvy, která nabývá na významu s množstvím klesajících srážek. Každoročně se zde vyskytují požáry, které vytváří selekční tlak ve prospěch ohnivzdorných druhů. Hustota stromové vrstvy záleží na půdních podmínkách a tlaku člověka. Otevřené formace se vyskytují ve formách od světlého savanového lesa přes stromovou, křovitou a travnatou savanu. Tyto různé typy savany lze nalézt i na malých plochách. Výskyt jednotlivých typů je závislý na fyzickogeografických podmínkách. V dnešní době je významná i činnost člověka – sběr dříví, spásání dobytkem atd. Na africkém kontinentu v současnosti převládají otevřené formace savan.
Světlé savanové lesy
Nejvyšší patra světlých savanových lesů tvoří opadavé stromy vysoké a střední velikosti (8-20 m). Koruny nevytváří zápoj. Mají široce rozložené koruny s malými listy. Dřevinný podrost se zde nevyskytuje. Bylinná vrstva je řídká, tvořená převážně vysokostébelnými trávami.
 |
| Ukázka světlého savanového lesa. (N.P. Kruger, Jižní Afrika) |
Flóra světlých savanových lesů
Flóra je tvořena jak tropickými rody deštných lesů (Afzelia, Brachystegia, Burkea, Daniella, Etythrophleum, Parinari, Uapaca), tak rody příbuznými (Isoberlinia, Psedoberlinia) a rody vyskytujícími se v lesích Guinejského zálivu (Anogeissus, Butyrospermum, Cochlospermum – J.-C. Menaut, 1992). V důsledku dlouhotrvajících požárů a čištění lesů jsou tyto lesy tvořeny většinou pouze jedním druhem stromů.
Komunita rostlin je přizpůsobena dlouhým obdobím sucha trvajícím 4 až 7 měsíců. Tento stres na životní prostředí vytváří několik charakteristických znaků světlých savanových lesů. Většina stromových druhů je opadavá během období sucha. Pupeny jsou chráněny před vysušením palisty nebo vlásky. Stromy jsou adaptovány na vysokou insolaci a teploty. Nevyskytují se zde epifyty, liánové druhy jsou vzácné. Suchá tráva a zaschlé výhonky snadno hoří a požáry zničí mnoho mladých výhonků. Požáry určují rozšíření a rozestupy stromů v savaně.
Savanové lesy se vyskytují v místech, kde se vyskytují dlouhá období sucha a srážky dosahují 650 až 1500 mm. Někdy se zde uplatňují i jiné klimatické podmínky. Např. súdánská provincie, která má vlhčí klima než province zambijská, má větší vodní deficit, protože zde vane od Sahary suchý vítr harmattan. Lesy súdánské provincie jsou proto více otevřené, méně druhově bohaté a méně vysoké než lesy zambijské provincie. Často se vyskytují na sušších a skalnatějších půdách než na jakých je okolní savana. V jižních částech Afriky je pás savanových lesů širší než na sever od rovníku. Některé savanové lesy nalezneme i na pobřeží Indického oceánu
Savanové lesy súdánské provincie
Zde byly zaznamenány tři kategorie savanového lesa. První je savanový les s dominantním rodem Isoberlinia, který se vyskytuje na rovinatých půdách. Na erodovaných svazích je nahrazen savanovým lesem tvořeným rodem Monotes a na chudých jílovitých půdách savanovým lesem Terminalia. Druhý je složením podobný prvnímu a není stále rozhodnuto o jeho platnosti. Třetím je savanový les s dominantním rodem Boswellia. Vyskytuje se na skalnatých podložích Senegalsko-súdánské oblasti.
Tyto otevřené lesy často přechází ve stromové savany s charakteristickými druhy stromů Daniella oliveri a Lophira lanceolata na jihu a Parkia biglobosa a Butyrospermum paradoxum na severu (J.-C. Menaut, 1992).
Savanové lesy oblasti východoafrických vysočin
Ve východním Kongu (Zaire) přežívají lesy pouze reliktně. Tvoří je druhy Albizia a Strychnos. Ve východní Africe se též vyskytují akáciové savanové lesy. Vyskytují se v údolích, na svazích a skalnatých plochách. V jižní části oblasti východoafrických vysočin nalezneme savanové lesy s rodem Brachystegia, zde nazývané miombo. Pokrývají značnou část Keni a dvě třetiny Tanzánie (J.-C. Menaut, 1992).
Savanové lesy zambijské provincie
Opět zde byly zaznamenány tři kategorie savanového lesa: savanový les s dominantním rodem Brachystegia pokrývá jižní části střední Afriky v nadmořských výškách okolo 1000 m n. m.. Druhým typem obývající podobné prostředí, ale v píscích Kalahari, je savanový les s dominantním rodem Baikiaea. Poslední kategorií je savanový les tvořený rodem Colophospermum, nazývaný také jako mopane. Snáší více suché podnebí a je rozšířen jižněji od dvou předcházejících kategorií.
Stromovité a keřovité savany
Na rozdíl od savanového lesa se stromovitá savana liší nižším zápojem, stromy se větví nížeji při povrchu. Floristicky se liší pouze rozdílnou skladbou dominantní vrstvy. Křovitá savana se od stromovité liší nižší výškou stromových druhů. Stromy mají dobře vyvinutý kmen a obvykle dosahují výšky 7 až 8 m. Keře mají malý a pokřivený kmen s nízko se oddělujícími se větvemi. Stromovité a křovité savany též pokrývají větší plochy než savanový les. Tvoří také přechod k travnaté savaně. V travnaté vrstvě stromy a keře rostou ve velkých rozestupech (viz. obr.5).
Stromovitá savana se vyskytuje převážně v západní Africe se stromovými druhy Butyrospermum paradoxum a Parkia biglobosa, v Kongu (Zaire) se stromovým druhem Cussonia angolensis. Křovitá savana je rozšířena po celé délce Sahelu s keři Hymenocerdia acida a Piliostigma thonningi. Ve většině křovitých savan Sahelu se vyskytují též stromy, které jsou vázány na stará termitiště.
Travnatá savana
V travnatých savanách ubývá podíl stromovité složky, jen občas se zde vyskytují solitérní stromy nebo keře. Ve skupinách se stromy a keře objevují pouze v místech, kde stará termitiště narušují lateritické slepence, a stromy tak mohou využívat zásob podzemní vody. Hlavní dominantní vrstvou je zde travnatý pokryv. Ten je výsledkem půdních podmínek (přírodní savany) nebo vznikl činností člověka (druhotné savany). V oblastech s vlhčím klimatem jsou výsledkem činnosti člověka.
 |
| Travnatá savana se stromovou složkou. V popředí antilopa vodní (Kobus ellipsiprymnus) (S. Luangwa N.P., Zambie) |
S druhotnými savanami se setkáme zvláště v oblastech tropického deštného lesa, které mohly v těchto oblastech vzniknout nejen činností člověka. Předpokládá se, že při klimatických změnách se savany šířily na jih. Laterizace změnila stavbu půdního horizontu a vytvořila tvrdé krusty. Deštný les se sem již nevrátil. Travnaté savany se též vyskytují na chudých píscích. Z trav zde dominuje Pennisetum purpureum nebo Imperata cylindrica. Tyto druhy též okupují místa ležící ladem. Travnaté savany s rodem Imperata se mohou snadno změnit zpět na stromovité savany. Stromy jsou často porostlé (např. Hyparrhenia rufa) a keři (Annona senegalensis, Bridelia ferruginea). Travnaté savany v pobřežních oblastech vznikly většinou v důsledku požárů a rychle se sem vrací savanový les.
Na rozšiřování travnatých savan se podílí i velcí savci, zvláště sloni (Loxodonta africana). Svou činností mění savanové lesy na travnaté savany. Tento úkaz byl zaznamenán hlavně na omezených plochách národních parků, kde svou činností negativně ovlivňuje biodiverzitu parků.
Přírodní travnaté savany se vyskytují v oblastech, kde délka období sucha poskytuje travám výhodu nad stromovými druhy. Dnes je známo množství rozličných kategorií závisejících na půdních a fyzických podmínkách.
 |
| Travnatá savana s křovitou složkou (Krugerův N.P., Jižní Afrika) |
Budeme-li postupovat od každoročně zamokřených savan do savan suchých, nalezneme zde toto schéma výskytu trav: Ve vlhkých loukách roste Vossia cuspidata a Oryza barthii, dále Echinochloa spp. lépe snášející střídání období dešťů a sucha. V oblastech s delším obdobím sucha porostou trávy Hyparrhenia rufa a Loudetia simplex, které vytváří jednolitý pokryv. V této zóně se vyskytují termitiště, nepoužívaná jsou porostlá stromovými druhy např. Acacia spp., Combretum spp.. Echinochloa roste po požárech ve vlhčích oblastech. Toto schéma je platné v oblasti střední Afriky a obecně se využívá pro celou oblast savany (J.-C. Menaut, 1992). Co se týče velikosti, v trvale podmáčených půdách a místech s dosažitelnou hladinou podzemní vody, dorůstají trávy obřích rozměrů. Mezi nimi vyniká Andropogon gayanus dosahující výšky až 6 m, zvaná „sloní tráva“. V druhotných savanách po zničeném tropickém sezónním lese roste až do výše 8 m. Mnohé jednoleté trávy dosahují výšky okolo 2 m, přestože mají většinou krátký vegetační cyklus trvající tři až čtyři měsíce. Opakem jsou trávy krátkostébelných savan, které dorůstají velikosti jen několika centimetrů (např. Microchloa kunthii a Sporobolus stapfianus). V oblastech s intenzivním spásáním a v lokalitách s krátkou dobou dešťů dorůstají trávy obecně menších rozměrů. Na místech s odlišnými vlastnostmi půdy se trávy v rámci jednoho druhů též značně liší. Např. druhy Andropogon canaliculatus, Schizachyrium schweinfurthii, Sprobolus pyramidalis a Vetiveria fulvibarvis dosahují v savanách kolem toků v guinejské a súdánské zóně Afriky výšky 2 m, kdežto v suchém okrsku mezi Togem a Ghanou jen výšky 1 m (Jeník, 1975). Rozdíly mezi šáchorovitými travinami jsou též značné a platí pro ně to samé, co pro trávy. Rozsáhlé plochy savan v okolí Viktoriina jezera pokrývá Loudetia kagerensis. V údolních oblastech jihozápadního Súdánu vytváří souvislou vrstvu trávy Hyparrhenia spp. a šedé jíly středního Súdánu pokrývá Brachiaria obtusiflora, neboť snáší i dlouhodobé zamokření.
 |
| Travnatá savana v každoročně zaplavovaném území. Stromovitá složka je vázána na vyvýšená místa, jako jsou termitiště. (Okavango delta, Botswana) |
V sušších oblastech se již vyskytuje nízkostébelná savana, kde jednoleté trávy dosahují menších velikostí než 80 cm. Nesplňují tak jednu z podmínek klasifikace biomu savan, vytvořenou koncilem v Yangambii. Hovoří se o nich jako o „stepích“ nebo „pseudo-stepích“, „pouštních savanách“ atd. Prof. Jeník pro ně nalezl výše jmenované označení – „nízkostébelná savana“. Splňují ale ostatní podmínky definice savany: výskyt dřevní složky, občasný výskyt požárů atd. V pásu Sahelu, při hranicích se Saharou, jsou nejčastější dřevní složkou Acacia radiata a Panicum turgidum. Tyto savany dále nalezneme v Kongu, Zairu a v guinejské zóně, která je sice klimaticky vlhká, ale v místech, kde půda neudrží vlhkost.
*Terofyty: jednoleté rostliny, prodělávající celý životní cyklus během jediného vegetačního období. Nepříznivou část roku přečkávají v semenech.
**Hemikryptofyty: vytrvalé nebo dvouleté rostliny, mající během nepříznivého období pupeny uložené jen při povrchu půdy, kryté živými nebo odumřelými listy, šupinami, pochvami listů apod.
***Fanerofyty: Převážně keře a stromy, popř. liány s obnovovacími orgány výše než 0,3 cm nad zemí. V této souvislosti k nim patří stromové epifyty, rostliny uchycené jen na povrchu fynerofyt nebo na nich parazitně či poloparazitně žijící.
****Geofyty: dvouleté až vytrvalé rostliny, přečkávající nepříznivé období pouze orgány v půdě hlouběji ukrytými (kořeny, oddenky, hlízami, cibulemi).
*****Chamaefyty: obnovovací pupeny mají nad povrchem půdy (nejvýše do 0,3 cm), chráněné v období vegetačního klidu jen šupinami nebo nahloučenými jinými nadzemními orgány.
Citace: Menaut J.-C.(1992): Ekosystems of the world – Tropical savanna, Elsevier, Amsterdam, s. 109-144
Jeník J. (1975): Život tropických savan II, Živa, 23, č. 2, Academia, Praha, s. 42-46
Chavalier (1933); Tchoumé (1966); Jacques-Félix (1959), Walter (1971) převzato z: Ekosystems of the world – Tropical savanna, Elsevier, Amsterdam, s. 109-144
4.5.Fauna
Africké savany jsou známy především díky početným populacím velkých savců. Přestože se ukázalo, že pro život a homeostázi ekosystému savan není pastevně kořistnická větev nezbytná (Jeník, 1975). Člověk procházející savanou však vnímá většinou pouze tyto velké savce, ptáky a obtížný hmyz. Malí živočichové a jiné životní formy, mající svou aktivitu v nočních hodinách nebo v období dešťů, zůstávají návštěvníkovi skryti.
Z hlediska zoogeografického členění patří africké savany do etiopské oblasti. Ta náleží do paleotropické říše. K etiopské oblasti přičleňujeme navíc jižní cíp arabského poloostrova, Madagaskar a ostatní ostrovy v blízkosti pobřeží.
Fauna etiopské oblasti je charakteristická značným výskytem endemických čeledí a podčeledí savců; z hmyzožravců např. zlatokrtovití (Chrysochloridae), bercounovití (Macroscelididdae), vydříkovití (Potamogalidae); z hlodavců jsou zde šupinatkovití (Anomaluridae), noháčovití (Pedetidae); z kopytníků žirafovití (Giraffidae), hrochovití (Hippopotamidae); dva řády jedné endemické čeledi – damani (Hyracoidae) a hrabáči (Tubulidentata); z ptáků turakovití (Musophagidae), hadilovovití (Sagittariidae) a kladivošovití (Scopidae).
Nejtěsnější vztahy etiopské oblasti lze prokázat s faunou indomalajské oblasti. Obě proto náleží do říše Paleotropis. V menší míře se uplatňují vztahy s palearktickou oblastí, v důsledku izolace během třetihor mořem Tethys.
Vnitřní členění etiopské oblasti není zřejmé, člení se na podoblasti jih, západ a východ, ale hranice jsou značně neostré. Naproti tomu je madagaskarská podoblast snadno definovatelná. Je to dáno její ostrovní izolací. Fauna ostrova se vyvinula z druhů, které sem migrovaly, převážně z afrického kontinentu. Vyvinuly se z nich samostatné čeledi. Přesto zde nežije mnoho taxonů typických pro africký kontinent: lichokopytníci (Perissodactyla), sudokopytníci (Artiodactyla), opice (Anthropoidae), myšovití (Muridae), jeřábi (Gruidae), dropi (Otididae), datlovití (Picidae), pěnkavovití (Fringillidae) atd. Pozoruhodné na madagaskarské fauně je její podobnost s říší Neotropis. Ze zmíněných důvodů (4.2.2.1.; Úvod), zde nebude Madagaskar uváděn.
Nejběžnějšími bezobratlými živočichy v savanách jsou členovci – pavouci a hmyz. Z členovců jsou časté různé druhy klíšťat a fytofágních roztočů, kteří obývají travnatou vrstvu savany. Hojní jsou též zástupci dvoukřídlých (Diptera) Drosophilidae a Calliphoridae. Dále do této skupiny patří komáři (rody Anopheles, Culex, Stegonia), přenášející původce onemocnění malárie (Plasmodium malariae) a moucha tse- tse, přenášející původce spavé nemoci (Trypanosoma brucei). Hojně rozšířené jsou mimo níže zmíněných všekazů i mravenci, vosy a včely. Pravděpodobně nejznámější z třídy hmyzu je v afrických savanách podřád sarančovití (Caelifera), tvořící v období sucha 40 až 50 % biomasy členovců. Je znám svými hromadnými přesuny. Přes všechnu škodu, kterou způsobují na obdělávané půdě, jsou významnou složkou potravy většiny živočichů savan. Hejna sarančat obsahují asi 700 milionů až 2 miliardy jedinců. Na jeden hektar pak připadá asi 2,5 tuny sarančat. Mezi „stěhovavé druhy“ africké savany patří saranče egyptská (Anacridium aegypticum), s. pustinná (Schistocerca gregaria) a s. stěhovavá (Locusta migratoria).
Termité jsou detritofágové. Rozmělňují a rozkládají odumřelou rostlinou hmotu v kořenové a povrchové vrstvě. Patří mezi nejpočetnější faunu savan. Termitů žije v savanách asi 600 druhů, někteří se živí výhradně suchou trávou, jiní dřevem, nebo pěstují houby. Všekaz bojovný (Bellicositermes bellicosus) má hustotu jedné stavby na 2 ha. Při stavbě termitiště je přesunuto 20 až 25 m3 zeminy. V termitišti žije populace od 200 000 až do milionu jedinců. Tato populace zkonzumuje až 50 % produkce travního krytu. Průměr živé váhy termitů na 1 ha je 5 kg (Jeník,1975). Jiný druh všekaze , všekaz mosambický (Horotermes mossambicus), je výjimka mezi termity. Neživí se pouze odumřelou hmotou, ale „sklízí“ i živé nadzemní části rostlin. Tento rod termita patří k primitivnějšímu stádiu vývoje termitů, nemá dokonale rozlišeny kasty a též způsob výživy není specializován.Termiti svou činností nepůsobí pouze na okolní rostlinstvo, ale mění i stavbu půdy a pozměňují reliéf. Na jejich opuštěných stavbách se často uchytávají stromy, protože je zde v dosahu podzemní voda načerpaná kvůli termoregulaci hnízda. Z jejich staveb buduje člověk silnice a dokonce mu slouží jako zdroj potravy. Termitiště se zde dědí z otce na syna. Potravou se stává i pro jiné živočichy, např. hrabáč kapský (Orycteropus afer) a hyenka hřivnatá (Proteles cristatus) se živí výhradně jimi. Termitiště slouží pro mnohé další živočichy jako místo úkrytu.
Velmi početní jsou též švábi (Blattidae) a cvrčci (Gryllidae). V ekosystému savany mají značný význam mravenci, přesto jsou však stále málo známí. Nejběžnějšími jsou omnivorní (všežravé) druhy, a dále druhy mravenců, kteří pěstují semena rostlin. Druhy karnivorní (masožravé) nejsou tak početné.
Žížalovití jsou nejdominantnější složkou půdní fauny. Hrají důležitou roli v asimilaci a mineralizaci organického materiálu. Jejich druhová rozmanitost, biomasa i aktivita je značná. Jejich činností vznikají v půdním profilu „kamenité čáry“, které vznikly přemisťováním jemnozrnného materiálu k povrchu.V oblasti Pobřeží Slonoviny žije na ploše 1 ha až 180 000 jedinců o živé hmotnosti 170 kg. Rozborem přítomných generací a odumřelých jedinců byla odhadnuta roční produkce živé váhy 460 kg na 1 ha (Jeník, 1975). Nejpočetnější složkou půdních členovců jsou roztoči. Přesto hustota členovců v půdě dosahuje asi jednu desetinu hodnoty udávané v Evropě (Jeník, 1975). Proč tomu tak je, není známo.
Ryby
Africkými savanami protéká množství vodních toků a rozkládají se zde četná jezera. Fauna primárně sladkovodních ryb je zde bohatá a charakteristická velkou přítomností archaických skupin (9 čeledí, z nichž je 7 endemických). Nejzajímavější z nich jsou bichirovití (Polypteridae) s 13 druhy. Řád dvojdyšných (Dipnoi), zastoupený v Africe bahníkem africkým, ukazuje na dřívější spojení Afriky s Jižní Amerikou a Austrálií. Největší podíl rybí fauny tvoří šest čeledí sumců s 250 druhy. V tropické oblasti Afriky nalezneme tři centra výskytu sladkovodních ryb. První a nejbohatší je povodí řeky Kongo. Dále reliktní jednotný systém Niger-Čad-Nil. Zcela nezávislým centrem výskytu sladkovodních ryb jsou východoafrická jezera. Dominuje v nich endemická čeleď (Cichlidae).
Obojživelníci
Z obojživelníků jsou zde zastoupeny především žáby (Anura). Ostatní ocasatí (Urodela) a červoři (Apoda) nejsou již tak běžní. Obojživelníci mají dvě životní fáze. První, larvální, žije ve vodním prostředí. Druhé, dospělec, po metamorfóze vylézá na souš. Jejich relativní nezávislost na vodním prostředí je narušena potřebou se množit. Jsou též velmi citliví k přeschnutí a potřebují alespoň vzdušnou vlhkost. Vyvinuli se proto u nich určité životní strategie, které možnému vysušení pokožky nebo narušení rozmnožovacího cyklu, předchází: Jsou aktivní pouze v podmínkách dovolujících jim přežít. Některé druhy jsou aktivní pouze v období dešťů. Jiné přesunuly svou aktivitu do nočních hodin, kdy je atmosférická vlhkost vzduchu vyšší než ve dne. Další žijí jen v blízkosti vod. Zajímavé jsou druhy žijící trvale pod zemí, ve vlhké a kypré půdě, jako například africký červoř Hemisinae. Právě jejich svázanost s vlhkem vysvětluje jejich rozšíření, sezónní změny v hustotě a v životním cyklu. Některé žáby se dokázaly přizpůsobit dennímu způsobu života. Mají tenkou epidermis s množstvím mukózních žláz. Vyšší sekrece slizu dovoluje regulovat teplotu skrz evaporaci pokožkou a ochlazovat tak povrch. Je to případ některých ropuch Bufo pentoni, B. xeros a některých druhů Tomoptera. Žába Pyxicephalus adspersus se naopak vyskytuje pouze v období dešťů (M. Lamotte, 1992). Mnoho druhů žab, které žijí v travách v okolí vodních toků a na stromech, má vyvinuto ochranné a rozptylující vzory. Druhy jako Hyperolius nasutus a H. lamottei žijí na dlouhých travách v blízkosti vod. Jsou velmi tenké a na zádech mají dlouhé tmavé pruhy. Stromové druhy mají jiné kryptické znaky. Barvu mají zelenou nebo světlehnědou a vzorem napodobují okolí nebo mají skvrny občas narušeny pruhy ( např. Kassina senegalensis, K. cassinoides) (M. Lamotte, 1992).
Plazi
Z plazů je v savanách relativně početný pouze řád Squamata (plazi šupinatí) z něhož převažuje čeleď Lacertidae – ještěrky. Krokodýli jsou vázáni na vodní prostředí a v Africe žije jen jeden druh – krokodýl nilský (Crocodylus niloticus). O želvách žijících v savaně není mnoho známo.
Hlavní rody plazů žijící v savanách jsou Mabuya a Agama z podřádu Sauria (ještěři) a Echis a Psammophis z podřádu Serpentes (hadi). Ostatní rody Squamat vyskytující se v Africe lze rozdělit podle následujících kategorií: kosmopolitní čeledi Colubridae (užovkovití) a Viveridae (zmijovití, nevyskytují se v Austrálii); pantropické čeledi Gekkonidae (gekonovití) a Scincidae (scinkovití) z řádu ještěrů a Typhlopidae (slepákovití), Boidae (hroznýšovití) a Elapidae (korálovcovití) z řádu hadů; a čeledi vyskytující se pouze v oblastech tropů Starého světa Agamidae (agamovití) a Chamaeleontidea (chameleonovití).
Rozmístění, velikost populací a rozšíření Squamat regulují záplavy v nížinných oblastech v době dešťů a požáry v biomech savanových lesů v době sucha. V místech, která jsou chráněna před účinky požárů v savanách, je dvakrát více Squamat než v okolní savaně. Během roku se tento rozdíl vyrovná migrací. K nejznámějším a pro člověka nejnebezpečnějším Squamatům Afriky patří mamba černá (Dendoaspis polylepis) a m. zelená (D. anguisticeps).
Ptáci
Bohatost ptačí fauny savan je větší než v deštném lese. Nalezneme zde 700 druhů ptáků z 57 čeledí. Je to dáno větší pestrostí biomu savany. C.H.Fry udává, že v savanových lesích s rodem Isoberlina žije 375 druhů na 10 000 km2. V savanách Senegalu lze spatřit 120 druhů na 25 ha. Nejpočetnější jsou snovači (Ploceidae) zastoupeny 61 druhy. Tito ptáci jsou zrnožraví. Další početnou paleotropickou čeledí je Estrildidae. Početní jsou též zrnožraví frankolíni (Phasianidae), hmyzožraví lejskové (Sylviidae). Poslední tři početné čeledi jsou orli (Aguilidae) s 33 druhy; strdimilové (Nectariniidae) s 23 druhy a vousáci (Capitonidae). Zajímavé jsou dvě endemické čeledi myšáci (Coliidae), živící se ovocem a Sagittariidea, zastoupená pouze jedním druhem.
Ptáci obývají především okolí savanových lesů a vodních toků. V době sucha je toto ještě více zvýrazněno. Jejich množství v savanách je větší než v tropickém deštném lese. V zimním období je ptačí fauna obohacena o stěhovavé druhy z Evropy. Charakteristickými většími ptáky savan jsou pštros dvouprstý (Struthio camelus), hadilov písař (Sagittarius serpentarius) a drop kori (Choriotis kori). Pštros je vůbec největší žijící pták světa. Pod nohami pasoucích se stád loví hmyz různé druhy volavek a čápů. Snovatec červenozobý (Quelea quelea) je pravděpodobně nejpočetnějším ptákem světa. Obývá suché savany Sahelu. Odhaduje se, že jeho populace čítá 1,5 miliardy jedinců. Jedna hnízdící kolonie může obsahovat až deset miliónů hnízd. Snovači pospolití si staví společné hnízdo obsahující až 250 hnízdních komor.
Savci
Z „velkých“ savců jsou v Africe početní různí kopytníci. Z čeledi turovití (Bovidae), mající asi 110 druhů se v afrických savanách vyskytuje 78 druhů. Řád Proboscidea je zastoupen jedním druhem – slonem africkým. Řád Perissodactyla (lichokopytníci) je v afrických savanách zastoupen 6 druhy. Řád Artiodactyla má 5 čeledí, které se zde vyskytují: čeleď Suidae s třemi druhy; Hippopotamidae s 2 druhy; Giraffidae s 2 druhy; Tragulidae s 1 druhem a výše zmíněnou čeleď Bovidae. Šelmy (Carnivora) jsou zmíněny v kap. 4.5.4..
Z „malých“ savců jsou zde zastoupeni myšovití (Muridae) s 79 druhy. Tito malí savci jsou typičtí r-stratégové a jsou zcela závislí na množství srážek, které určují úrodu travních semen. Dalšími četnými čeleděmi řádu hlodavců (Rodentia) jsou: veverkovití (Sciuridae); křečci (Cricetinae); plši (Muscardinidae); řekomyšovití (Thryonomyidae); hlodouni (Rhizomyinae); dikobrazi (Hystricidae); noháčovití (Pedetidae) a rypošovití (Bathyergidae). Početní jsou i zajíci rodu Lepus, kaloni a netopýři. Většina známých velkých savců afrických savan je zmíněna v kapitole 4.5.4..
Velké množství býložravých zvířat savany využívá toto prostředí rozličným způsobem. Živí se různě vysokými rostlinnými patry. Malí živočichové, jako jsou hlodavci, mají k dispozici čerstvě rostoucí kořeny, hlízy, listí a plody. Podzemní kořeny a hlízy vyrývají prasata bradavičnatá (Phacochoerus africanus). Sloni a žirafy dokáží spásat listí vysoko v korunách stromů. Listí nižších stromů a keřů slouží za potravu nosorožcům dvourohým (Diceros bicornis), antilopě losí (Taurotragus oryx), kudu (Tragelaphus) a na zadní nohy se stavějící antilopě žirafí (Litocranius walleri). Větévky jen několik centimetrů nad zemí okusují antilopky travní (Raphicerus campestris) a dikdici (Madoqua).
Býložravci živící se převážně trávou taktéž využívají tento potravní zdroj různými způsoby. Svou spotřebu energie kryjí různým výběrem pastvy a různou strategií zažívání. Nepřežvýkavci využívají potravu méně intenzivně a proto nejsou při jejím výběru moc vybíraví, např. hroši, sloni a zebry. Přežvýkavci nejsou schopní pozřít tak velké objemy potravy. Proto věnují větší pozornost výběru pastvy, patří mezi ně buvoli, antilopy, gazely, buvolci a pakoni. Při migracích za zdroji potravy nastupují do porostů trav v tomto pořadí. První se pasou zebry (Equus sp.) spásající vrcholky trav, buvoli afričtí (Synceros caffer), pakoni bělobradí (Connochaetes taurinus) a buvolci (Damaliscus sp.) je následují. Za potravu jim slouží střední listnatá část trav. Nejnižší a nejšťavnatější část nebo čerstvě rašící rostliny spásají antilopy. Jako představitele posledně jmenovaných uveďme gazelu Thomsonovu (Gazella thomsoni), gazelu Grantovou (Gazella granti), antilopu skákavou (Antidorcas marsupialis) a impalu (Aepyceros melampus).
Uprostřed pasoucích se stád antilop loví nejznámější africká šelma, lev (Panthera leo). Žije ve smečkách. Loví hlavně samice. Na kořist útočí ze zálohy. Kořistí se mohou stát i tak velcí býložravci, jako je buvol. V afrických savanách loví ještě dvě další kočkovité šelmy, gepard (Acinonyx jubatus) a levhart (Panthera pardus). Gepardi pronásledují zpravidla menší kořist rychlými sprinty. Levharti útočí z úkrytů v blízkosti napajedel. Pes hyenovitý (Lycaon pictus) loví ve smečkách a kořist pronásleduje tak dlouho, dokud ji neuštve. Z menších šelem lovících především hlodavce a bezobratlé zde nalezneme mangusty, cibetky, promyky, surikaty a žanetky z čeledi cibetkovitých (Viverridae). Z čeledi kunovitých se zde vyskytuje medojed (Mellivora capensis), který je znám ze soužití s ptákem medozvěstkou (Indicator indicator). Tento pták se snaží svým výstředním chování a zvukem přilákat medojeda k hnízdu včel. Medozvěstka pak ze zničeného hnízda vybere larvy včel.
Ze šelem se mršinami živí hyeny. Nejznámější je hyena skvrnitá (Crocuta crocuta), charakteristická svým „smíchem“. Na mršinách se též živí čáp marabu (Leptophilus crumeniferus), supi bělohlaví (Gyps fulvus), s. hnědí (Aegypius monachus) a šakal čabrakový (Canis mesomelas), jinak lovící malé obratlovce.
Paviáni (Papio) žijí ve skupinách a jejich potravou jsou kromě kořenů, hlíz a bobulí i drobní obratlovci a hmyz. Patří tedy do kategorie všežravců.
Tyto potravní vazby jsou jen velmi zjednodušeným potravním řetězcem, ukazujícím pouze „velké“ savanové živočichy. Vztahy producentů, primárních a sekundárních konzumentů jsou ve skutečnosti mnohem složitější. Vše, co v každém ekosystému končí dentrito-saprofágní větví zde nebylo podrobně zmíněno.
") ")  usedá na větve s nohama ve výši lopatek (Vumba, Zimbabwe)")  je nejnápadnějším ptákem savany (Kruger N.P., Jižní Afrika)") ") (Kruger N.P., Jižní Afrika)")  patří k nejběžnějšímu druhu ledňáčka v Africe (Okavango delta, Botswana)")  dorůstá výšky až 5,7 metrů a je často lovena pro svou oháňku, ze které se vyrábí náramky (Chobe N.P., Botswana)") ") ") ") ") ") ") |
| Popis obrázků dostanete přidržením myši nad obrázkem |
Citace: Fry C.H. (1992): Ekosystems of the world – Tropical savanna, Elsevier, Amsterdam, s. 337-354
Lamotte M. (1992): Ekosystems of the world – Tropical savanna, Elsevier, Amsterdam, s. 313-322
Jeník J. (1975): Život tropických savan IV., Živa, 23, č. 5, Academia, Praha, s. 163-167
4.6. Oheň v savaně
Podle definice přijaté vědeckým koncilem pro Afriku na mítinku fytogeografie v Yangambii je savana:“útvar trav vysokých alespoň 80 cm, který tvoří dominantní vrstvu nižších trav, obvykle každoročně vypalovanou požáry. Trávy mají ploché listí, bazální metabolismus a stonky. Obvykle jsou zde zastoupeny stromovité rostliny.“ (koncil Scientifique pour l´Afrique, 1956). Někdy se hovoří dokonce o tom, že savany jsou ohňový-klimax („fire-climax“). Požáry mají velký vliv na půdu, vegetaci i faunu savany.
Požáry jsou častější v místech s většími srážkami, protože přírůstky biomasy trav jsou v těchto místech mnohem větší než v sušších oblastech. Během období sucha poskytují tedy více materiálu, který shoří během každoročně se opakujících požárů. Požáry savany vznikají přírodní cestou (savana je zapálena bleskem nebo tlením a následnou pyrolýzou rostlinného materiálu nad 50°C ), nebo jsou založeny člověkem. Člověk zjistil, že savanové trávy se po požárech rychleji obnovují a poskytují příhodnější pastvu pro jeho dobytek, ohně jsou používány k lovu nebo k zakládání ploch pro zemědělství. Vypalování trav v Africe člověkem je staré asi 50 000 let (D. Gillon, 1992). Požáry v savaně tedy většinou zakládá člověk, zvláště v oblastech, kde má lidská populace větší hustotu.
Růst teploty během požárů savany nemá dlouhého trvání. Teplota roste v okolí ohňové fronty jen asi pět až šest minut. Nejvyšší teploty trvají jen několik sekund. Teplota při povrchu půdy se liší místo od místa. Dosahuje velikosti mezi 75 až 350°C. Nejvyšších teplot je dosaženo v travách vysokých 1,2 až 1,5 m. Teplota v 50 cm nad zemí dosahuje maximálně 500 až 600°C. Požáry na začátku období sucha zničí přibližně 25 % travního pokryvu, ale na konci toto množství vzrůstá. Pozdní požáry savany zničí 64 až 96 % travní vrstvy (D. Gillon, 1992). Požáry v počátku období sucha jsou mnohem šetrnější pro stromové porosty a k půdě. Z hlediska zemědělství jsou naopak výhodnější požáry na konci období sucha, protože zničí více plevelu a náletových stromů. V oblastech národních parků se proto doporučuje vypalování ihned na začátku období sucha. Jak ukázaly výzkumy na plochách chráněných před požáry se travnatá savana pomalu mění ve stromovitou nebo křovitou. Je však nesprávné z tohoto vývoje činit závěr, že savana s dřevinami je vždycky přirozeným závěrem sukcese daného místa (J. Jeník, 1976).
Nejvíce hořlavou vrstvou je množství suchého listí. Dřevnaté části rostlin hoří jen výjimečně. Listí trav je bohaté na různé proteiny, fosfor, vápník a dusík. Požáry tak výrazně urychlují koloběh živin v savanách. I když je nutné podotknout, že kromě dusíku unikajícího do atmosféry (okolo 70 %), se ztrácí i část jiných živin, které jsou odnášeny větrem v podobě jemných částic.
Jak bylo výše zmíněno požáry mají velký vliv na půdu savan. Tím jak spalují půdní pokryv vystavují půdu účinkům deště a slunečnímu záření. Zvyšuje se tak půdní eroze. Záleží však též i na topografických vlastnostech terénu.
Vliv ohňů na půdní živočichy je různý. Výskyt ohňů snižuje druhovou bohatost u pavouků, roztočů a vidličnatky Diplura, které jsou zvláště početné. Naopak některým živočichům každoroční ohně vyhovují, např. humusem se živícím žížalám, larvám brouků, termitům „sklízečům semen“ atd..
Oheň má též zničující účinek na obojživelníky, plazi a obecně pro všechny, kteří nemají dost schopností se před ohněm skrýt nebo utéct. Přesto byly zaznamenány i úhyny velkých savců, jako je slon nebo lev. V populacích velkých savců padnou za obět požárům pouze mladí jedinci nebo slabé, nemocné a zraněné kusy. Největší užitek poskytují požáry ptákům, kteří jsou chráněni před následky, díky schopnosti letu. Ptáci a jiní živočichové vítají ohně v savaně jako snadný prostředek opatřit si potravu. Před a nad frontou požáru poletují kukačky, vlaštovky, rorýsy a různí krahujcovití. Ty následují mandelíci, vlhy a volavky (např. Ardeola ibis). Po průchodu ohně přilétají čápi marabu (Leptoptilos crumeniferus) a vrány (Corvus albus). Společně s ptáky na obětech požárů hodují hyeny a šakalové. Pár týdnů po požáru vypadá savana jako bez života. Po vyrašení prvních výhonků trav se začínají objevovat býložravci, které následují jejich predátoři.

Požáry v savanách jsou zakládány převážně lidmi, tráva se po přechodu ohně rychle obnovuje a poskytuje lepší pastvu pro dobytek a divokou zvěř (Kafue N.P:, Zambie)
Citace: Gillon D. (1992): Ekosystems of the world – Tropical savanna, Elsevier, 1. vydání, Amsterdam, s. 617-638
Jeník J. (1976): Život tropických savan I., Živa, 24, č.1, Akademia, Praha, s. 2-5
4.7. Vliv člověka
Hlavním ekologickým činitelem v afrických savanách je člověk. Svou činností ovlivňuje vegetaci, faunu, půdu a vodní režim. V dnešní době převyšuje populace člověka všechny ostatní živočichy. Kromě parazitů nemá člověk v savaně přirozeného nepřítele. Proto je nutné se o úloze člověka v savanách zmínit.
Původ člověka je dnes hledán v afrických savanách. Savany jsou spjaty s člověkem od počátku jeho vývoje. Předci člověka nacházely v savanách lepší životní prostředí než v tropickém deštném lese. Celý lidský vývoj byl proto determinován prostředím savany. Lidské smysly, vzpřímená postava, psychika i jeho sociální chování jsou adaptace na biom savan. Každé z těchto přizpůsobení pomáhalo člověku přežít. I v dnešní době jsou savany nejhustěji osídleny.
Díky různorodosti savan lze zde zaznamenat všechny stupně vývoje lidské společnosti. Od lovců sběračů přes pastevectví, rolnictví až po moderní zemědělství. V některých oblastech se tvář savan změnila natolik, že zde nenalezneme žádné zbytky přírodní krajiny, ale pouze plně industriální lidskou společnost. Hlavním faktorem v přetrvávání různých stupňů vývoje společnosti jsou podmínky fyzickogeografické. Nejdůležitějším limitním faktorem je potřeba vody. V suchých oblastech přetrvává společnost lovců a sběračů nebo kočujících pastevců, v blízkosti vodních ploch se rozmohlo rolnictví a moderní zemědělství. Hlavní křižovatkou v tomto vývoji se stala východoafrická jezera.
Lovci a sběrači
Nejznámější příklady lovců a sběračů na africkém kontinentu jsou Křováci a Pygmejové. Druzí jmenovaní obývají pouze deštné lesy. Křováci neboli Sanové dnes žijí pouze v oblastech polopouště Kalahari. Dříve byli rozšířeni po celém regionu jižní Afriky. Nejprve byli zatlačováni khoikhoiskými pastevci (Hotentoti), kteří se stěhovali po západním pobřeží Afriky. Později se k nim přidaly bantuské kmeny od severovýchodu. Poslední ránu jim zasadil příchod bílých osadníků. Osadníci vyhubili při osídlování jižních oblastí všechnu zvěř, a tak byli Sanové nuceni lovit búrský dobytek. Búrové během osmi let zabili na dva a půl tisíce Sanů. Búrů přišlo o život asi dvě stě. I doktor Emil Holub nazval tuto část historie osídlování země jako „nejohyzdnější skvrnu v dějinách Kapska“. Sanové tak byli nuceni se stáhnout do těchto nehostinných oblastí. Přesto se dokázali těmto podmínkám znamenitě přizpůsobit. Vodu Sanové získávají z občasných vodních ploch, dužnatých okurkovitých plodů i jiných rostlin. Za potravu jim dále slouží hlízy rostlin, vejce pštrosů, ještěrky, žáby, mravenci, všekazi a kobylky. Dokáží ulovit i větší zvěř pomocí otrávených šípů. Jsou též výborní stopaři a dokáží se k zvěři přiblížit do těsného kontaktu, využívajíc nerovností terénu, napodobováním tvarů lovné zvěře a jejich zvuků. Křováci si opatřují potravu ve skupinách. Žijí ve společenstvích dvaceti až sta lidí. Větší skupiny tvoří jen v období sucha, kdy se soustřeďují okolo zdrojů vody. Křováci mají i určitou formu soukromého vlastnictví. Skupina, která žije na určitém území si označuje plodné rostliny a termitiště. V dnešní době se vláda Botswany, na jejímž území žije drtivá populace Sanů, snaží tyto kočující kmeny přimět k usedlému životu. Jinou cestou se vydali v Jižní Africe, kde letošního roku poskytli Sanům k životu podle jejich tradic národní park Kalahari Gemsbok. Přesto i zde budou pod civilizačním tlakem. Na oplátku za poskytnuté území budou plnit roli stopařů zvěře pro turisty. Podle odhadů žije tradičním životem v oblasti Kalahari jen 2000 Sanů.
Kočovní pastevci
Kočovní pastevci obývají převážně travnaté savany. Stáda skotu potřebují neustálý přísun potravy. Tráva je v těchto oblastech velmi sporá v důsledku malých srážek. To nutí tyto kmeny neustále migrovat. Ke kočovným pastevcům patří některé kmeny Bantů, Fulbů a turisticky nejvíce navštěvovaný kmen Masajů. Tento způsob života byl umožněn domestikací skotu, ovcí a koz. Společnosti pastevců jsou většinou patriarchální. S ochranou stád skotu je u nich patrna i určitá bojovnost a nadřazenost vůči usedlým rolníkům. Např. Masajové si podrobily kmen Wandorobů, který pro ně zabíjí domácí zvířata mimo skotu. Opatřují si u nich i plodiny, které sami nepěstují. V rozdělení kmenů na usedlé zemědělce a kočující pastevce lze hledat příčinu krvavého konfliktu mezi Huty a Tutsii ve Rwandě. Vzájemná symbióza mezi člověkem a jeho stády dosáhla vrcholu u Masajů. Masajové nepotřebují k obživě většinou nic jiného než krev skotu a mléko. Maso jedí pouze z uhynulých kusů dobytka. Tato silná specializace se najde jen u některých národů. Většinou je chov dobytka kombinován s pěstováním kulturních plodin.
Rolníci
Rozvoj lidské populace v afrických savanách nastal s pěstováním obilnin a okopanin. Teprve objev rolnictví umožnil růst lidské populace a na příhodných místech i vznik velkých civilizací. Důležitým předpokladem pro rolnictví je vhodný typ půdy a dostatek vláhy pro kulturní plodiny. Půda savan má vysoký obsah vápníku, hořčíku a někdy i solí. Pod travnatou vrstvou savan je příznivé množství humusu. Ten však poskytuje užitek jen omezenou dobu, protože se vlivem vypalování a přeměny savany v ornou půdu rychle rozkládá. Nepříznivým faktorem pro zemědělství je i výskyt lateritických slepenců, které omezují zásoby půdní vláhy. Střídání období dešťů a sucha vyžaduje též vhodné plodiny, které stačí dozrát v takto limitovaných podmínkách. Z tradičních obilnin jsou pěstovány různé druhy čiroků a prosa (např. rody Echinochloa, Eleusine, Digitaria, Panicum, Paspalum a Pennisetum). Zrno se drtí pro výrobu mouky, slouží k výrobě tradičních piv nebo se používá jako krmivo pro drůbež. Některé druhy prosa snáší i velmi nepříznivé klimatické podmínky a rostou i v oblastech s velmi malými srážkami. Dále se pěstují různé druhy čiroků (rod Sorghum). Z dalších plodin mají význam kořenové hlízy jamů (Dioscorea sp.), hlízy manioku, ibišek jedlý a různé druhy čeledi okurkovitých (Citrullus vulgaris, Cucumis sativum a Lagenaria siceraria). Významnou plodinou je i podzemnice olejná (Arachis hypogea). Ze stromovitých rostlin je hospodářsky využíván baobab (Adonsonia digitata).
Moderní zemědělství
S objevitelskými cestami se k africkým zemědělcům dostaly i plodiny, které zde nemají svůj původ. K nejznámějším patří kukuřice a pšenice. Pšenice je pěstována jen v oblastech s dostatečnými srážkovými poměry. V místech, kde je možné umělé zavlažování se pěstuje rýže. Naopak kukuřice je dnes velmi rozšířenou kulturní plodinou, zvláště v jižní Africe. Tyto tři obilniny úspěšně vytlačují původní tradiční plodiny. Mají větší výnosnost z hektaru a jsou chutnější. Přes poměrně příznivé fyzickogeografické podmínky se v Africe nepovedla „zelená revoluce“. Tento experiment zde zkrachoval na tradičním kmenovém zřízení, nedostatečné infrastruktuře a značné chudobě afrických státu. Proto zde nedošlo k většímu využívání mechanizace a chemizace v zemědělství. V oblasti Sahelu k tomu pomáhají i značně nerovnoměrné srážky s opakujícími se výskyty velkých such. Pokusy domestifikovat některé africké kopytníky ztroskotaly (buvol kaferský, antilopa losí). Úspěšně však byli domestifikováni pštros dvouprstý a částečně i antilopa skákavá.
Industriální společnost
V místech s výskytem nerostných surovin africká savana ustoupila tlaku člověka. Vyrostla zde velká města a průmyslové závody. S nimi dochází k rozvoji dopravy. Nová spojení zpřístupňují dříve odlehlé oblasti vlivu člověka, který narušuje ekologickou stabilitu těchto míst. Devastace savan v Africe probíhá podle stejného vzorce, jakého bylo uplatněno v listnatých a jehličnatých lesích západní Evropy.
Z obecného pohledu devastace přírodního prostředí nastává ve chvíli, kdy si člověk začíná přizpůsobovat krajinu k obrazu svému. Rozvoj lidské společnosti, bohužel, vždy přináší určité poškozování prostředí ve kterém žije. Pouze lovci a sběrači žijí v souladu s přírodou. Jejich společnost je determinována prostředím, ve kterém žijí. S objevem zemědělství byl započat proces přeměny přírodní krajiny v kulturní. Vhodné podmínky savan na africkém kontinentu předurčily proces zkulturňování tohoto biomu více než deštného lesa nebo polopouštních oblastí. První zemědělci zabírali jen malé plochy savan. Množství parazitů a obtížného hmyzu v blízkosti vodních toků a málo výnosné plodiny nedovolovaly větší rozvoj lidské populace. S domestifikací skotu, koz a ovcí nastaly první problémy. Člověk potřeboval pastviny pro svá stáda, která konkurovala divoké zvěři. Přesto následky pro životní prostředí nebyly velké. Teprve s příchodem moderní civilizace nastaly vážné problémy. S příchodem bílých kolonizátorů nastalo vykořisťování divoké přírody. Rozmohl se obchod se slonovinou. Později začaly osadníci zabírat plochy savan pro svůj dobytek a při tom decimovaly populace afrických kopytníků. Taktéž stáda domorodých pastevců poškozovaly krajinu nadměrným spásáním. Povrch půdy se úplně obnaží a nejúrodnější svrchní vrstva půdy je vystavena erozi. Dnes se lze setkat i se situací, kdy majetní občané si dodnes drží stáda skotu jen pro znamení moci. Jejich bohatství je počítáno podle kusů dobytka. S příchodem výnosnějších druhů obilovin a zlepšením zdravotní péče nastal prudký růst lidské populace. Člověk zabírá stále větší plochy savan. To je průvodní jev extenzivního zemědělství. Naproti tomu na růst intenzifikace zemědělství nemají africké státy finanční prostředky, navíc je trh zaplaven levnějšími zemědělskými produkty z jiných částí světa. Divoká zvěř je tak zatlačována na území národních parků. Navíc původní obyvatelé Afriky jsou vášniví lovci a mimo národní parky se s velkou zvěří nelze setkat. Nejhorší dopady na životní prostředí savan nastávají v okamžiku, kdy se k činnosti člověka přidají i podmínky klimatické.
Nejlepším příkladem toho je rozšiřování Sahary na úkor travnatých savan sahelské zóny. Ze skalních kreseb v pohoří Tassili lze usuzovat, že desertifikace potkala i celou oblast Sahary. Na nejstarších skalních kresbách (okolo 6 000 př.n.l.) jsou zobrazována savanová zvířata (hroši, žirafy, sloni a antilopy). Okolo roku 5000 př.n.l. se na skalních kresbách již objevují stáda skotu. Jedno století před naším letopočtem jsou zde znázorněni již pouze jezdci na velbloudech. Otázkou zůstává jestli se tento výrazný přechod savany v poušť neodehrával pouze za značného přispění člověka.
Pokračující desertifikace sahelské zóny je zapříčiněna periodicky se vyskytujícím delším obdobím sucha. Sucho sebou nese neúrodu trav a kulturních plodin. Dochází k úhynu dobytka a posléze i k hladomoru v lidské populaci. Lidskou tragédii doprovází i tragédie biomu savan. Příroda je za normálních okolností k déle trvajícímu suchu adaptována. Po příchodu dešťů se travní kryt nejrychleji obnovuje v okolí křovin a stromů, které ve svém okolí zadržují travní semena. Lidé však v těchto oblastech potřebují k přípravě pokrmů palivové dříví, které proto těží. Jiný zdroj paliva se v těchto oblastech nevyskytuje. Savana během suchých let tyto ztráty nemůže nahradit. Vše je umocněno velikostí lidské populace. Tím jak dochází k odstranění stromové vegetace, půdní kryt tak není ničím chráněn. Do krajiny vstupuje vítr a odnáší svrchní část půdy. Během několika let sucha je v oblastech s dřívějším výskytem savany v lepším případě polopouštní step. Tento závažný proces degradace krajiny je bezpochyby mnohem složitější než je zde popsáno. Vyřešit tento problém nebude jednoduché. Nejúčinnějším řešením by bylo opětovné zalesnění krajiny i za cenu introdukce nepůvodních stromových druhů.
") . V blízkosti lidských sídel rostou též banánovníky a mangovníky (Chisenga, Zambie)")  je nejrozšířenější obilninou savanových oblastí jižní Afriky (Kalongola, Zambie)") |
| Popis obrázků dostanete přidržením myši nad obrázkem |
Citace: Holub E. (1890): Druhá cesta po Jižní Africe, 1. díl, nakladatelství J.Otto, Praha
4.8. Ochrana přírody a národní parky
Africké savany byly a jsou pod neustálým tlakem člověka. Nejvíce trpí populace velkých savců jejichž stavy se neustále snižují. Proto byly již na počátku minulého století zakládány v savanách velkoplošné rezervace a národní parky. Prvním byla velkoplošná rezervace Sabi, později přejmenovaná na Krügerův N.P. (založena roku 1898). Poté bylo založeno i mnoho jiných rezervací a národních parků. V dnešní době nalezneme v Africe asi 700 chráněných území o ploše 1,34 milionů km2. Z tohoto počtu je zde asi 126 národních parků (mapa 9). Plošně největším je národní park Kafue mající rozlohu 2,2 tisíc km2. Plošně největší jsou však rezervace v pouštních oblastech, např. Ouadi Rimé/Ouadi Achim v Čadu a rezervace Air a Téréné v Nigeru (obě okolo 80 tis. km2). Důležitou roli v ochraně přírody zde hrají i soukromé rezervace. Patrně nejcennějším chráněným územím v Africe jsou národní parky Albertův a Královny Alžběty. Každý z nich leží na území jiného státu, ale dohromady vytváří dvojpark. Vyhnul se tak klasické chybě ostatních chráněných území, kdy ochrana přírody je limitována hranicemi států. K nejznámějším parkům patří národní park Serengeti, známý každoročními migracemi kopytníků. Tohoto jevu si pravidelně všímají i světová média. Zahynou zde totiž v rozdivočelých vodách řeky Mary tisíce pakoňů bělovousích (Connochaetes taurinus). Podobně bohatým parkem na kopytníky je méně známý park Liuwa Plain.
Plošně omezená chráněná území přinesla i nečekané problémy. Ochrana je soustředěna většinou na velké obratlovce. Zákaz střílení a ochrana před pytláky způsobily nárůst populací těchto savců. Populace zvěře přestaly migrovat. Navíc se na území parků přesouvá i zvěř z jiných oblastí, protože zde není obtěžována lovci a pytláky. Rezervace nejsou schopny však tyto populace uživit a to vede k nadměrnému tlaku na vegetaci a půdu. Nejlépe zdokumentována je v tomto směru činnost slonů. Za příklad může sloužit národní park Murchisonovy vodopády a Chobe. První z nich se dříve rozkládal ve stromovité savaně. Avšak během třiceti let se stromová savana změnila v travnatou. Sloni svou činností (lámání větví, loupání kůry a vyvracení stromů) prosvětlují zapojené lesnaté porosty. Na ploše parku žije asi 8 000 slonů, kteří s každoročními požáry dokončují takto započatou proměnu (F. Bourliére, 1992).
Národní park Chobe je z hlediska rozmanitosti typů savan bohatší. V současné době zde žije 70 tisíc slonů. Předpokládaná zátěž populace slonů na prostředí je jen poloviční. Jediným řešením je regulovaný odstřel. Ten však naráží na odpor celosvětové veřejnosti. V současné době se proto v Krügerově národním parku zabývají studií, jak by bylo možno slony sterilizovat. Jiným řešením je slony nebo jinou zvěř bezpečně odchytit a přetransportovat na území, ve kterých byly dříve vyhubeni.
Úspěšnými projekty byla např. introdukce nosorožce tuponosého (Ceratotherium sinum) v některých parcích na území Jižní Afriky a Zimbabwe.
Jiné problémy vznikají při zakládání a správě parků s místními obyvateli. Například národní park Serengeti musel být zmenšen oproti původnímu návrhu. V kráteru Ngorongoro se podle dohody mezi vládou a Masaji pase okolo 2000 kusů dobytka. Největší problémy jsou však v nejchudších afrických zemích. Místní obyvatele nutí nedostatek potravy pytlačit v chráněných oblastech. Horší variantou je lov trofejové zvěře. Např. finanční obnos pytláka za roh nosorožce vystačí lovci na celý rok. Dalším vážným problémem je těžba palivového dříví.
Zajímavým projektem, bohužel zatím myšlenkovým, je vytvoření tzv. „Afrického superparku“ (mapa 8). Projekt byl vypracován pro region jižní Afriky. Současné parky a rezervace by se měli propojit soustavou biokoridorů nebo zvětšením stávající plochy. Prvním projektem by mělo být propojení třech národních parků: Krügerova, Banhine a St. Lucia.
Vznikem „afrického superparku“ by bylo možno obnovit staré migrační cesty, které tvoří systém riftových údolí a pohoří severojižního směru, doplněný příčně tekoucími řekami k Indickému oceánu. Obří park by měl napomoci vedle zvýšení počtu divoce žijící zvěře, k rozšíření a obohacení genofondu. Výhody jsou spatřovány v integrovaném přístupu v ochraně přírodního prostředí, stejně tak v turistickém managementu i v kontrole pytláctví. Turističtí experti přesvědčují, že takovýto park by vedl nejen k ochraně ohrožených druhů živočichů, ale pozvedl by ekonomiku afrických zemí na přijatelnou bázi (Kliment, 1998).
Dříve byly parky zakládány bez znalosti ekologických vazeb a pouze k ochraně velkých savců, v případě uskutečnění tohoto projektu by se napravily chyby z minulosti.
Citace: Kliment Z. (1998): Africký superpark – sen nebo realita, Geografické rozhledy, 97-98, č.4, Česká geografická společnost, Praha, s. 115
Bourliére F. (1992): Ekosystems of the world- Tropical savanna, Elseviere, Amsterdam, s. 730
Stuart Ch. a T. (1997): The large Mammals of Afrika, Struik, Cape Town, s. 319
5. Závěr
Smyslem práce bylo podat co nejúplnější charakteristiku biomu savan. V díle jsou zachyceny hlavní fyzickogeografické podmínky, jako jsou podmínky klimatické, geologické a pedologické, ovlivňující rozšíření savan v Africe. Podnebí je charakterizováno střídáním období sucha a dešťů, na které je vázán hlavní vegetační růst.
Značná pozornost byla věnována savanové vegetaci, na které jsou přímo či nepřímo, závislí všichni živočichové. Vegetace též v závislosti na fyzickogeografických podmínkách určuje ráz (fyziognomii) savany. Dominantní životní formou jsou početné druhy trav a šáchorovitých, které soutěží s dřevinami o využití půdní vlhkosti a živin v půdě. Savana vykazuje značnou proměnlivost, variabilitu v prostoru a čase. Tato proměnlivost je tedy jejím nejdůležitějším znakem. Vyskytuje se ve formách přecházejících z tropického lesa až po otevřenou step v tropické zóně.
Kapitola „Oheň v savaně“ vysvětluje důležitost požárů v ekosystému savan. Požáry jsou spolu s býložravci, živinami v půdě a vlastnostmi půdy hlavními modifikačními faktory. Tato kapitola však neospravedlňuje nešetrné počínání v zakládání požárů lidmi.
Nemalou měrou byla věnována pozornost úloze člověka v savaně. Mnoho primatologů a antropologů spatřuje počátky našeho druhu v savanách, kde první prohominidé přešli ze stromů do otevřené savany. Savana je jakýsi „skanzen“ lidské populace. Byla patrně osídlena lidmi již v počátcích jejich vývoje. Role člověka v savaně je dnes natolik významná, že v některých místech nelze rozlišit jestli je savana přírodního nebo druhotného původu. V některých oblastech došlo též k nevratnému poškození tohoto biomu. Naštěstí si místní lidé začali uvědomovat význam savan pro život jejich i jiných organismů. Proto na územích jednotlivých států začaly vznikat chráněné rezervace a národní parky. Kapitola o vlivu člověka na savanu spíše pokládá více otázek, než na které se snažila odpovídat a zasloužila by si vlastní zpracování.
Savany jsou důležitým místem ekologické proměnlivosti a mají klíčovou roli v evoluci biosféry a mnoha druhů, ze kterých se skládá. Savany představují mladý biom v geologickém měřítku. Nabídly některým tropickým organismům nové příležitosti. Ty se úspěšně adaptovaly daným podmínkám životního prostředí. Jakékoli rychlé a inovační zásahy proto značně zatěžují prostředí africké savany.
") ") ") ") ")   |
| Popis obrázků dostanete přidržením myši nad obrázkem |
6. Seznam literatury
Adámek H., Hanzelka J., Zikmund M. (1996): Živá Afrika, Trango, Praha, s. 175
Attenborough D. (1990): Planeta žije, Panorama, Praha, 334 s.
Bourliére F. (editor) (1992): Ekosystems of the world, 13 – Tropical savanna, Elseviere, Amsterdam, s. 730
Buchar J. (1983): Zoogeografie, SPN, 1. vydání, Praha, s.199
Dorst J. (1974): Ohrožená příroda, Orbis, Praha, s. 406
Ellis W. S. (1987): Africa´s Stricken Sahel, National Geographic, vol. 172, č. 2, NGS, Washigton, D.C., s. 140-179
Few R., Duncan B., Campbell S, Zahrniser E. (1995): Atlas panenských míst, Knižní klub, Praha, s. 240
Grove A.T. (1978): Afrika, 3. vydání, Oxford UN. Press, Oxford
Grundelingh A. a kol. (1996): South Afrika yearbook, Sable media, CapeTown, s. 542
Hendrych R. (1983): Fytogeografie, SPN, 1. vydání, Praha, s. 220
Holub E. (1890): Druhá cesta po Jižní Africe, 1. díl, nakladatelství J.Otto, Praha, s. 576
Holub E. (1890): Druhá cesta po Jižní Africe, 2. díl, nakladatelství J.Otto, Praha, s. 608
Horník S. a kol. (1986): Fyzická geografie II, SPN, 1.vydání, Praha, s. 319
Chadwick D. H. (1995): Ndoki last place on Earth, National Geographic, vol. 188, č. 1, NGS, Washigton, D.C., s. 2-45
Jeník J. (1995): Ekosystémy (Úvod do organizace zonálních a azonálních biomů), 1. vydání, skripta, Karolinum, Praha, s. 135
Jeník J. (1991): Život tropických savan I až VI, Interní xerox – Živa, PřF UK, Praha, přerušované strany
Jeník J. (1994): The Dahomey gap, Mém. Soc. Biogéogr., 3. série, s. 125-133
Jeník J. (1975): Život tropických savan I, Živa, 23, č. 1, Academia, Praha, s. 2-5
Jeník J. (1975): Život tropických savan II, Živa, 23, č. 2, Academia, Praha, s. 42-46
Jeník J. (1975): Život tropických savan III, Živa, 23, č. 3, Academia, Praha, s. 84-88
Jeník J. (1975): Život tropických savan IV, Živa, 23, č. 5, Academia, Praha, s. 163-167
Jeník J. (1975): Život tropických savan V, Živa, 23, č. 6, Academia, Praha, s. 202-205
Jeník J. (1976): Život tropických savan VI, Živa, 24, č. 1, Academia, Praha, s. 2-5
Kliment Z. (1994): Problém zvaný Sahel, Geografické rozhledy, 4, č. 1 (94-95), ČGS, Praha, s. 18-19
Kliment Z. (1997): Viktoriiny vodopády, Geografické rozhledy, 7, č. 3 (97-98), ČGS, Praha, s. 77-78
Kliment Z. (1998): Africký superpark – sen nebo realita, Geografické rozhledy, 7, č. 4 (97-98), ČGS, Praha, s. 114-116
Knapp D. (1973): Die Vegetation von Afrika, 3. vydání, G. Fisher, Jena, s. 626
Kol. autorů (1994): Obratlovci, 1. vydání, Nakladatelský dům OP, Praha, s. 687
Kubeš J. (1996): Jižní Afrika – základní informace, Velvyslanectví Jihoafrické republiky v Praze, Praha, s. 133
Kunský J. (1935): Všeobecný zeměpis, 1. díl, knihkupectví R. Prombergra, Olomouc, s. 396
Kunský J. Málek R., Vrána O. (1971): Zeměpis světa- Afrika, 1. vydání, Orbis, Praha, s. 546
Lhote H. (1987): Oasis of Art in the Sahara, National Geographic, vol. 172, č. 2, NGS, Washigton, D.C., s. 180-191
Maclean G. L. (1993): Roberts´ Birds of Southern Africa, John Voelcker Bird Book Fund, 6. vydání, Cape Town, s. 871
Netopil R. a kol. (1984): Fyzická geografie I, SPN, 1.vydání, Praha, s. 272
Obrtel R. (1983): Království zvířat, 1. Vydání, Albatros, Praha, s.136
Payne K. (1989): Elephant talk, National Geographic, vol. 176, číslo 2, NGS, Washigton, D.C., s. 262-277
Rosypal S. (1987): Přehled biologie, 1. vydání, SPN, Praha, s. 686
Steinerová J. (editor) (1995): Savany – Larousse život v přírodě, Knižní klub, Praha, s. 166
Stuart Chris a Tilde (1997): The larger mammals of Africa, Struik, 1. vydání, Cape Town, s. 319
Theroux P (1997): Zambezi, National Geographic, vol. 192, číslo 4, NGS, Washigton, D.C., s. 2-31
Vágner J. (1978): Afrika-ráj a peklo zvířat, Svoboda, Praha, s. 205
Vágner J. (1979): Afrika-život a smrt zvířat, Svoboda, Praha, s. 241
Zeman J. (1991): Africký Kaleidoskop, Mladá fronta, Praha, s. 165
7. Sezman obrázků, map a tabulek
Obr.1 Schéma vazeb po odstranění dobytčího moru s.15
Obr.2 Slon africký, foto Chobe N. P., Botswana s.16
Obr.3 Železitá krusta, foto Ngonye falls,Zambie s.36
Obr.4 Typy savan v Africe s.38
Obr.5 Baobab, foto Nata, Botswana s.42
Obr.6 Akácie, foto Okavango delta, Botswana s.51
Obr.7 Stálezelený les, foto Krügerův N.P., Jižní Afrika s.55
Obr.8 Světlý savanový les, foto Krügerův N.P., Jižní Afrika s.57
Obr.9 Travnatá savana se stromovou složkou, foto S. Luangwa N.P., Zambie s.60
Obr.10 Travnatá savana s křovitou složkou, foto Krügerův N.P., Jižní Afrika s.61
Obr.11 Travnatá savana, foto Okavango delta, Botswana s.62
Obr.12 Kladivoš, foto Okavango delta, Botswana s.64
Obr.13 Africká kobylka, foto Lobatse, Botswana s.66
Obr.14 Čáp marabu, foto Krügerův N.P., Jižní Afrika s.70
Obr.15 Spárák šedý, foto Krügerův N.P., Jižní Afrika s.70
Obr.16 Myšák pruhovaný, foto Vumba, Zimbabwe s.70
Obr.17 Ledňáček Ceryle rudis, foto Okavango delta, Botswana s.71
Obr.18 Mandelík fialovoprsý, foto Krügerův N.P.,Jižní Afrika s.71
Obr.19 Žirafy, foto Chobe N.P., Botswana s.72
Obr.20 Antilopka travní, foto Krügerův N.P., Jižní Afrika s.73
Obr.21 Zebra Böhmova, foto S. Luangwa N.P., Zambie s.74
Obr.22 Šakal čabrakový, foto Krügerův N.P., Jižní Afrika s.75
Obr.23 Hyena skvrnitá, foto Krügerův N.P., Jižní Afrika s.76
Obr.24 Lev, S. Luangwa N.P., Zambie s.76
Obr.25 Pavián babuin, foto Krügerův N.P., Jižní Afrika s.77
Obr.26 Požár v savaně, foto N.P. Kafue, Zambie s.79
Obr.27 Skot, foto západní Zambie, Sioma s.83
Obr.28 Maniok, foto Chisenga, Zambie s.84
Obr.29 Zemědělská usedlost, foto Kalongola, Zambie s.85
Obr.30 Pakoni a zebry, foto Krügerův N.P., Jižní Afrika s.88
Obr.31 Sloni u řeky Elephans´river , foto Krügerův N.P., Jižní Afrika s.89
Obr.32 Nosorožci, foto Matopos N.P., Zimbabwe s.90
Obr.33 Zebry u umělého napajedla, foto Krügerův N.P., Jižní Afrika s.91
Obr.34 Hroši, foto N. P. Kafue, Zambie s.91
Mapa 1; Mapa rozšíření savan ve světě s.19
Mapa 2; Průměrné roční srážky na africkém kontinentě s.26
Mapa 3; Nejvyšší průměrné srážky během nejvlhčích šesti měsíců na africkém kontinentě s.27
Mapa 4; Průměrné srážky během třech nejsušších měsíců na africkém kontinentě s.28
Mapa 5; Průměrné roční teploty na africkém kontinentě s.29
Mapa 6; Průměrná nejnižší teplota nejchladnějšího měsíce na africkém kontinentu s.30
Mapa 7; Mapa hlavních vegetačních typů africké savany s.39
Mapa 8; Mapa projektu afrického „superparku“ zpracovaného pro jižní Afriku s.92
Mapa 9; Národní parky Afriky s.93
Tab.1 Vegetační kategorie afrických savan
Tab.2 Procentuální zastoupení různých životních forem rostlinných druhů v některých afrických botanických oblastech

